يلعب الدوبامين (دا) العصبي دورا أساسيا في مختلف الوظائف المعرفية والسلوكية، ويتورط خلل وظيفته في العديد من الأمراض والاضطرابات الشائعة. على هذا النحو، فمن الأهمية بمكان لتطوير طرق دقيقة لدراسة كمية دنا نقل العصبي في الجسم الحي لتقييم كيفية تغيير العصب دا في سياقات نماذج المرض وعلم الأدوية المخدرات. سريع المسح الضوئي فولتامتري دوري (فسكف) يسمح للرصد في الجسم الحي دا العصبي مع قرار المكاني والزماني غرامة. في حين أنه من الممكن لمراقبة العصبي دا الفسيولوجية في مستيقظا، الحيوانات تتصرف بحرية، والتحفيز الكهربائي من مسارات الدوبامين تصاعدي في الحيوانات تخدير يمكن أن تنتج ردود دا قوية التي هي قابلة للتحليل الحركي تعزيز دا نقل العصبي.
تعكس ردود فعل دا المحفزة كهربائيا تفاعلا ديناميكيا لإطلاق سراح دا وإعادة امتصاصه، والتفسيراتمن هذه الردود قد استخدمت في الغالب نموذج بسيط من حفز دا العصبي دعا ميكلايس-منتن (مم) نموذج 12 . ويتألف نموذج مم من 3 متغيرات لوصف استجابات دا من حيث معدل إطلاق ثابت دا وكفاءة إعادة امتصاص ثابتة ( أي العلاقة بين معدل امتصاص دا وتركيزات دا خارج الخلية)، كما هو موضح في المعادلة 1 : 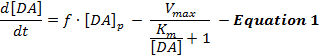
(الإفراج دا) (دا امتصاص)
في المعادلة 1، و هو تردد التحفيز. [دا] p هو زيادة تركيز دا المقدرة لكل نبض من التحفيز. V ماكس يمثل معدل امتصاص القصوى المقدرة. و K m هو ثابت مم المقدرة، وهو ما يعادل نظريا لتركيز خارج الخلية دا التي تشبع 50٪ من دات، مما يؤدي إلى معدل امتصاص نصف القصوى. هذا الاختلافيمكن دمج معادلة آل لمحاكاة ردود دا التجريبية بتقدير معلمات [دا] p و V ماكس و K m .
على الرغم من أن نموذج مم قد سهلت تقدما كبيرا في فهم حركية الناقل العصبي دا في سياقات تجريبية مختلفة، فإن نموذج مم يجعل الافتراضات الأساسية التبسيط التي تحد من انطباقه عند نمذجة ردود دا التي أثارها التحفيز سوبرافيسيولوجيكال 2 ، 13 . على سبيل المثال، يمكن لنموذج مم تقريب أشكال الاستجابة دا فقط إذا كانت ترتفع بطريقة محدبة، ولكنها لا يمكن أن تمثل ردود فعل تدريجية (مقعرة) المتصاعدة الموجودة في المناطق الظهرية المخططة 12 . وبالتالي، فإن افتراضات نموذج مم لا تعبر بدقة عن إطلاق ديناميكية وعمليات إعادة امتصاص الناقلات دا المحفزة.
لنموذج استجابات دا حفز وفقا لكمية واقعيةالإطار الإطاري، تم تطوير الإطار الكهربي العصبي الكمي (قن) استنادا إلى مبادئ حركية العصب الحركي المستمدة من البحث التكميلي والتجريب 2 . وتظهر خطوط مختلفة من البحوث العصبية أن (1) تحفيز الافراج العصبي هو عملية ديناميكية التي تنخفض في معدل على مدى التحفيز 14 ، (2) لا يزال الافراج في مرحلة ما بعد التحفيز مع حركية الاضمحلال ثنائي الطور 15 ، و (3) دا تمنع كفاءة امتصاص تدريجيا خلال مدة التحفيز نفسه 2 ، 16 . هذه المفاهيم الثلاثة هي الأساس لإطار قن، والمعادلات الثلاثة تتكون من 12 معلمات تصف ديناميات الإفراج دا وإعادة امتصاص ( الجدول 1 ). يمكن لإطار كن محاكاة عن كثب أنواع التجاوب التجريبي دا غير المتجانسة، وكذلك pالآثار المصابة للتلاعب التجريبية من المعلمات التحفيز وإدارة المخدرات 2 ، 6 . على الرغم من أن مزيد من البحث هو ضروري لصقل نهج النمذجة البيانات، يمكن للتجارب في المستقبل الاستفادة إلى حد كبير من هذا النهج النمذجة ترتكز على نيوروبيولوجيكالي، مما يضيف إلى حد كبير إلى الاستدلالات المستمدة من نموذج دا الناقل العصبي حفز.

الجدول 1: معادلات النمذجة والمعلمات . الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
يصف هذا البرنامج التعليمي كيفية نموذج حفز بيانات الاستجابة دا لتقدير دا إطلاق وحركية ريوبتيك باستخدام قنسم 1.0. جمع البيانات التجريبية الفعلية والعلاقات العامةلم يتم وصف أوسيسينغ هنا ويتطلب فقط بيانات تركيز دا الزمنية. وقد تم وصف الدعم النظري والأسس لإطار قن بشكل موسع سابقا 2 ، ولكن يرد وصف للمنظور العملي لتطبيق إطار الشبكة القطرية لنموذج بيانات استجابة جدول أعمال التنمية أدناه.
الإطار القاري نموذج التفاعل الديناميكي بين: 1) دينامية إصدار دا، 2) دا امتصاص، و 3) آثار التحفيز الفوقي السلفي على هذه العمليات لاستخراج المعلومات الحركية ذات مغزى من بيانات الاستجابة دا. إطار قن هو الأنسب لنمذجة فسكف البيانات المكتسبة باستخدام التحفيز فوق الفسيولوجية للغاية لفترة طويلة (على سبيل المثال، 60 هرتز، 10 ثانية التحفيز)، والتي تنتج ردود دا قوية التي هي قابلة للتحليل الحركي. وبعد النمذجة الدقيقة لعمليات الإطلاق وإعادة الامتصاص الأساسية، يمكن استعمال معلمات النموذج لمحاكاة استجابة دا التي ينبغي أن تقارب شكل المقياس السابقاستجابة تجريبية دا.
وتصف معادلات إطار الشبكة القطرية معدلات إطلاق سراح دا وإعادة امتصاصها على مدى ردود دا المحفزة. يصف إطار كن معدل إطلاق دا المحفز كدالة من الوقت من بداية التحفيز (t)، عندما ينخفض معدل إطلاق دا بشكل كبير على مدى التحفيز. ويتسق ذلك مع استنفاد تجمع قابل للنشر بسهولة، مع إضافة معدل إطلاق دا ثابت الحالة (دارس) لحساب تجديد الحويصلات، على غرار التقارير الأخرى ( المعادلة 2 ) 14 ، 17 .
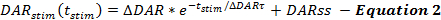
يؤدي التلاعب الذي يزيد من معدل إطلاق دا، مثل زيادة Δ دار أو Δ دار أو دارس، إلى زيادة اتساع الاستجابة على دا مقابل المؤامرات الزمنية. كل بارامتر يساهم بشكل تفاضلي في أشكال الاستجابة دا. زيادة دارس و Δ دار τ على حد سواء جعل المرحلة الصاعدة من الردود أكثر خطية (أقل محدبة). تناقص Δ دار τ يعزز التحدب، الذي يسيطر عليه مقدار Δ دار. استنادا إلى تجربة النمذجة، دارس عموما أقل من 1/5 من Δ دار؛ وبالتالي، Δ دار هي معلمة الافراج التي تحدد في المقام الأول اتساع الاستجابة الشاملة للاستجابة دا.
يتم وضع نموذج معدل تحفيز دا بعد التحفيز من خلال المعادلة 3 كاستمرار لمعدل إطلاق دا حفز من نهاية التحفيز (دار إس ) كدالة من الوقت بعد التحفيز (ر آخر ). يتبع معدل إطلاق دا بعد التحفيز نمط تسوس ثنائي الطور، كما هو موضح سابقا 15 ، مع مرحلة تسوس الأسي السريع ومرحلة تسوس خطية لفترات طويلة لنموذج اثنين كاعمليات الإفراج العصبي التي تعتمد على لسيوم.
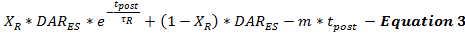
(التسوس الأسي السريع) (الاضمحلال الخطي المطول)
ليس من الممكن حاليا لتحديد مدى ما بعد التحفيز إصدار دا يحدث. ويمكن معالجة هذا الحد عن طريق التقليل المنهجي من تقديرات ما بعد التحفيز إصدار دا والتحقق من صحة المعلمات نموذج عبر مجموعة من الردود دا التجريبية التي تم جمعها من نفس الموقع تسجيل باستخدام فترات التحفيز متفاوتة. يسمح هذا التقليل للمستخدمين بإجراء تقديرات متحفظة من الإفراج وإعادة امتصاص. لأن التحفيز الكهربائي يؤدي إلى تراكم الكالسيوم الذي يعزز بعد تحفيز الناقلات العصبية الإفراج، ومدة التحفيز يؤثر على ما بعد التحفيز العصبيونميتر ريليس باراميترز 18 ، 19 . واستنادا إلى تجربة النمذجة، وجد أنه كلما زادت مدة التحفيز، يزداد τ R وينخفض X R ، بما يتفق مع الآثار المتوقعة لزيادة تراكم الكالسيوم 20 .
وتصف المعادلة 4 معدل امتصاص دا باعتباره امتدادا لإطار مم ويشتمل على المدى K ديناميكية، مما يزيد خلال التحفيز لنموذج تدريجيا انخفاض كفاءة امتصاص الناجمة عن التحفيز سوبرافيسيولوجيكال 2 ، 16 . يتم الاحتفاظ K م بعد التحفيز ثابتة في قيمة K م في نهاية التحفيز (K ميس).
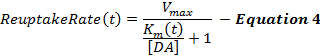
أين،
<img alt = "المعادلة 6" سرك = "/ فيليز / ftp_upload / 55595 / 55595eq6.jpg" />
(خلال التحفيز) (بعد التحفيز)
وغالبا ما تكون ردود الفعل المحفزة من دا، وخاصة من المناطق القبلية البطنية، غير حساسة للتغيرات في قيمة K m الأولية (كي مي )، الأمر الذي يجعل تحديد قيمة K مي مشكلة. وهكذا، مثل إطار مم الأصلي، يتم تقريب K مي في 0.1-0.4 μ M لاستجابات دا جمعها من السيطرة على الحيوانات غير المعالجة 12 . و Δ K المدى المدى يحدد مدى تغيير كفاءة امتصاص خلال التحفيز، والتي من تجربتنا حوالي 2081. M على مدى 60 هرتز، 10-ق التحفيز. وتحدد قيم k و مينف كيف تتغير K m بمرور الوقت، ويؤدي أي من هذين المصطلحين إلى تعزيز تقعر المرحلة الصاعدة. V ماكس هو معدل امتصاص القصوى التي تتصل جزئيا لكثافة دا الناقل المحلي، الذي يعرض فينتروميديال إلى التدرج الظهراني الوحشي 21 . وفقا لذلك، القيم V ماكس في المخطط الظهري (D- ستر) هي عموما أكبر من 30 μ م / ث ولكن عموما أقل من 30 μ م / ث في المناطق البطني، مثل نواة أكومبنس (ناك) 6 .
ويمكن للمبادئ التوجيهية العامة أعلاه أن تساعد في نمذجة بيانات الاستجابة التجريبية دا، ولكن توليد محاكاة تقارب استجابة دا التجريبية يتطلب تعديل نموذجي المعلمات نموذجيا. دقة المعلمات نموذج يمكن تحسينها عن طريق الحصول على ردود دا للتحفيز سوبرافيسيولوجيكال التي تقدمركيزة قوية للمحاكاة، وكذلك من خلال الحصول على نماذج متعددة دا والاستجابة لها لتحفيز فترات متفاوتة في نفس موقع التسجيل (على سبيل المثال، 60 هرتز، 5-S و 10-S التحفيز) للتحقق من صحة المعلمات ( انظر بيانات العينة). للتدليل، يتم تضمين مجموعة البيانات مع حزمة البرامج التي تحتوي على ريجيوسبيسيفيك تحفيز ردود دا جمعت في نواة أكومبنس والمخطط الظهري، قبل وبعد التحدي الدوائي الذي تم بالفعل نمذجة باستخدام إطار قن. في النهاية، سوف تجد المستخدمين يمكن تطبيق هذه المنهجية بالمثل لتوصيف حركية العصبي دا في سياقات المرض المختلفة والتلاعب الدوائية.
