La neurotransmission de la dopamine (DA) joue un rôle essentiel dans diverses fonctions cognitives et comportementales, et son dysfonctionnement est impliqué dans plusieurs maladies et troubles courants. En tant que tel, il est essentiel de développer des méthodes précises pour étudier quantitativement la neurotransmission de DA in vivo pour évaluer comment la neurotransmission de DA est modifiée dans le contexte des modèles de maladie et de la pharmacologie des médicaments. La voltamétrie cyclique à balayage rapide (FSCV) permet de surveiller la neurotransmission DA in vivo avec une résolution spatiale et temporelle fine. Bien qu'il soit possible de surveiller la neurotransmission de DA physiologique chez des animaux éveillés et qui se comportent librement, la stimulation électrique des voies dopaminergiques ascendantes chez les animaux anesthésiés peut produire des réponses de DA robustes qui sont susceptibles d'une analyse cinétique améliorée de la neurotransmission de DA.
Les réponses de DA stimulées électriquement reflètent un jeu dynamique de la version et du recapitulation de DA et des interprétationsDe ces réponses ont utilisé principalement un modèle simple de neurotransmission de DA stimulée appelée le modèle de Michaelis-Menten (MM) 12 . Le modèle MM se compose de 3 variables pour décrire les réponses de DA en termes de taux de libération de DA constant et d'efficacité de recapture constante ( c.-à-d., La relation entre le taux de recaptage de DA et les concentrations de DA extracellulaires), comme décrit par l'équation 1 : 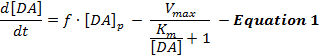
(Version DA) (recouvrement de DA)
Dans l'équation 1, f est la fréquence de la stimulation; [DA] p est l'augmentation estimée de la concentration de DA par impulsion de stimulation; V max représente le taux maximal estimé de recapture; Et K m est la constante estimée de MM, qui est théoriquement équivalente à la concentration de DA extracellulaire qui sature 50% de DAT, ce qui entraîne un taux de recaptage maximal. Cette différenceL'équation égale peut être intégrée pour simuler les réponses expérimentales de DA en estimant les paramètres [DA] p , V max et K m .
Bien que le modèle MM ait facilité des progrès significatifs dans la compréhension de la cinétique de neurotransmission de DA dans divers contextes expérimentaux, le modèle MM constitue des hypothèses fondamentales simplistes qui limitent son applicabilité lors de la modélisation des réponses de DA provoquées par des stimulations supraphysiologiques 2 , 13 . Par exemple, le modèle MM ne peut qu'approximer les formes de réponse DA si elles augmentent de manière convexe, mais elles ne peuvent pas expliquer les réponses progressives (concaves) croissantes trouvées dans les régions striatales dorsales 12 . Ainsi, les hypothèses du modèle MM ne capturent pas précisément les processus dynamiques de libération et de recapture de la neurotransmission de DA stimulée.
Modéliser les réponses de DA stimulées selon un quantum réalisteLe cadre quantitatif neurobiologique (QN) a été développé sur la base des principes de la cinétique de neurotransmission stimulée provenant de recherches et d'expérimentations complémentaires 2 . Différentes lignes de recherche sur les neurotransmissions démontrent que (1) la libération de neurotransmetteurs stimulée est un processus dynamique qui diminue dans le taux de stimulation 14 , (2) la libération se poursuit dans la phase post-stimulation avec la cinétique de décroissance biphasique 15 et (3) DA L'efficacité de la recapture est progressivement inhibée pendant la durée de la stimulation elle-même 2 , 16 . Ces trois concepts servent de base au cadre QN, et les trois équations se composent de 12 paramètres décrivant la dynamique de la libération et du recouvrement de DA ( tableau 1 ). Le cadre QN peut simuler de manière étroite des types de réponse expérimentale expérimentale hétérogènes, ainsi que la pEffets redacceptés des manipulations expérimentales des paramètres de stimulation et de l'administration du médicament 2 , 6 . Bien que des recherches supplémentaires soient nécessaires pour affiner l'approche de la modélisation des données, les expériences futures peuvent grandement bénéficier de cette approche de modélisation neurobiologiquement fondée, ce qui ajoute de manière significative aux inférences tirées du paradigme de neurotransmission de DA stimulé.

Tableau 1: Modélisation des équations et des paramètres . Cliquez ici pour voir une version plus grande de ce chiffre.
Ce didacticiel décrit comment modeler les données de réponse DA stimulées pour estimer la libération de DA et la cinétique de recapture en utilisant QNsim 1.0. La collecte de données expérimentales réelle et prLe traitement n'est pas décrit ici et nécessite uniquement des données temporelles de concentration de DA. Le support théorique et les fondements du cadre QN ont été largement décrits précédemment 2 , mais une perspective pratique sur l'application du cadre QN aux données de réponse DA modèle est décrite ci-dessous.
Le cadre de QN modélise l'interaction dynamique entre: 1) la libération de DA dynamique, 2) la recapture de DA et 3) les effets des stimulations supraphysiologiques sur ces processus pour extraire des informations de cinétique significatives à partir des données de réponse de DA. Le cadre QN est le mieux adapté à la modélisation des données FSCV acquises en utilisant des stimulations hautement supraphysiologiques de longue durée ( p. Ex., 60 Hz, 10 s stimulations), qui produisent des réponses de DA robustes susceptibles d'analyse cinétique. Suite à la modélisation précise des processus de libération et de recouvrement sous-jacents, les paramètres du modèle peuvent être utilisés pour simuler une réponse DA qui devrait se rapprocher de la forme de l'exRéponse expérimentale de DA.
Les équations du cadre de QN décrivent les taux de libération et de recouvrement de DA au cours des réponses de DA stimulées. Le cadre QN décrit le taux de libération de DA stimulé en fonction du temps depuis le début de la stimulation (t stim ), lorsque le taux de libération de DA diminue de façon exponentielle au cours de la stimulation. Ceci est conforme à l'épuisement d'un pool facilement libérable, avec un taux de libération de DA (DARss) à l'état stationnaire supplémentaire pour tenir compte de la reconstitution des vésicules, similaire à d'autres rapports ( équation 2 ) 14 , 17 .
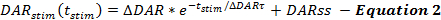
Les manipulations qui augmentent le taux de libération de DA, telles que l'augmentation de Δ DAR, Δ DAR τ ou DARss, conduisent à des amplitudes de réponse accrues sur DA contre les tracés temporels. Chaque parameTer contribue différemment aux formes de réponse de DA. Augmenter DARss et Δ DAR τ rendent la phase croissante des réponses plus linéaire (moins convexe). Diminuer Δ DAR τ favorise la convexité, qui est contrôlée par la grandeur de Δ DAR. Sur la base de l'expérience de modélisation, DARss est généralement inférieur au 1/5 de Δ DAR; Donc, Δ DAR est le paramètre de libération qui détermine principalement l'amplitude de réponse globale d'une réponse DA.
Le taux de libération de DA post-stimulation est modélisé par l'équation 3 comme continuation du taux de libération de DA stimulé à partir de la fin de la stimulation (DAR ES ) en fonction du temps après la stimulation (t post ). Le taux de libération de post-stimulation DA suit un schéma de décroissance biphasique, comme décrit précédemment 15 , avec une phase de décroissance exponentielle rapide et une phase de désintégration linéaire prolongée pour modéliser deux caProcessus de libération de neurotransmetteurs dépendants du calcium.
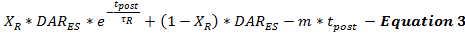
(Dégradation exponentielle rapide) (Dégradation linéaire prolongée)
Il n'est actuellement pas possible de déterminer la quantité de publication post-stimulation DA. Cette limitation peut être résolue en minimisant de manière systématique les estimations de la publication DA de post-stimulation et en validant les paramètres du modèle à travers un ensemble de réponses de DA expérimentales collectées à partir du même site d'enregistrement à l'aide de différentes durées de stimulation. Cette minimisation permet aux utilisateurs de faire des estimations prudentes de la libération et du recouvrement. Parce que les stimulations électriques conduisent à l'accumulation de calcium qui favorise la libération des neurotransmetteurs post-stimulation, la durée de la stimulation influe sur les neurotrans post-stimulationMitter release parameters 18 , 19 . Sur la base de l'expérience de la modélisation, on a constaté qu'à mesure que la durée de stimulation augmente, τ R augmente et X R diminue, ce qui correspond aux effets prévus d'une accumulation accrue de calcium 20 .
L'équation 4 décrit le taux de recaptage de DA comme une extension du cadre de MM et incorpore un terme de K m dynamique, qui augmente pendant la stimulation pour modéliser une efficacité de recapture progressivement décroissante causée par les stimulations supraphysiologiques 2 , 16 . Le K m après la stimulation est maintenu constant à la valeur K m à la fin de la stimulation (K mES ).
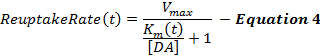
où,
<img alt = "Equation 6" src = "/ files / ftp_upload / 55595 / 55595eq6.jpg" />
(Pendant la stimulation) (après stimulation)
Les réponses stimulées de DA, en particulier des régions striatées ventrales, sont souvent insensibles aux changements de la valeur initiale de K m (K mi ), ce qui rend la définition d'une problématique de K mi . Ainsi, comme le cadre MM original, K mi est approximativement à 0,1-0,4 μ M pour les réponses DA collectées à partir des animaux témoins non traités 12 . Le terme Δ K m détermine l'ampleur du changement d'efficacité du recapture pendant la stimulation, qui, de notre expérience, est d'environ 2081; M au cours d'une stimulation de 60 Hz, 10 s. Les valeurs k et K minf déterminent comment K m change au fil du temps et l'augmentation de ces deux termes favorise la concavité de la phase ascendante. V max est le taux de recapture maximal qui concerne en partie la densité locale du transporteur DA, qui présente un gradient ventromédial à dorsolatéral 21 . En conséquence, les valeurs de V max dans le striatum dorsal (D-Str) sont généralement supérieures à 30 μ M / s mais généralement inférieures à 30 μ M / s dans les régions ventrales, comme le noyau accumbens (NAc) 6 .
Les directives générales ci-dessus peuvent aider à modéliser les données expérimentales de réponse DA, mais générer une simulation qui se rapproche de la réponse DA expérimentale nécessite un ajustement itératif des paramètres du modèle. La précision des paramètres du modèle peut être améliorée en obtenant des réponses DA à des stimulations supraphysiologiques qui fournissentUn substrat solide pour la simulation, ainsi que l'obtention et la modélisation de multiples réponses de DA à des stimulations de différentes durées sur le même site d'enregistrement ( p. Ex., 60-Hz, 5-s et 10-s stimulations) pour valider la précision des paramètres ( Voir les exemples de données). Pour démontrer, un ensemble de données est inclus avec le progiciel contenant des réponses DA stimulées par régiospécifique recueillies dans le noyau accumbens et le striatum dorsal, avant et après un défi pharmacologique qui a déjà été modélisé en utilisant le cadre QN. Par extension, les utilisateurs trouveront que cette méthodologie peut également être appliquée pour caractériser la cinétique de neurotransmission de DA dans divers contextes de maladie et des manipulations pharmacologiques.
