Durante el último siglo, la cristalografía de rayos X ha sido fundamental para dilucidar y comprender el paradigma estructura-función de las macromoléculas biológicas. Hasta la fecha, sigue siendo uno de los métodos más exitosos para dilucidar las estructuras de resolución atómica de muchas proteínas singularmente diferentes que son cruciales para la comprensión fundamental de la bioquímica celular, la medicina y el descubrimiento temprano de fármacos 1,2. Sin embargo, la cristalización de proteínas sigue siendo un cuello de botella en el estudio de muchas dianas proteicas, particularmente proteínas de membrana y grandes complejos proteicos3. En consecuencia, la cristalización de proteínas casi siempre se considera un arte debido a los enfoques de ensayo y error intensivos en mano de obra empleados 4,5,6.
Por lo general, se agrega un agente precipitante a una solución de proteína a alta concentración para formar una disposición reticular bien ordenada, regular y repetitiva de moléculas de proteína, conocidas como cristales. En condiciones favorables, como temperatura, pH, concentración y agente precipitante, finalmente se forma una solución sobresaturada, seguida de nucleación cristalina y crecimiento 7,8. Aunque ha habido muchos avances en las configuraciones de prueba de cristalización, predominantemente con el desarrollo de sistemas robóticos de alto rendimiento y la disponibilidad de pantallas de “matriz dispersa” listas para usar, los enfoques generales para la cristalización de proteínas se han mantenido sin cambios a lo largo de los años. Las técnicas experimentales comunes de cristalización de proteínas incluyen la difusión de vapor (gota colgante y gota sentada)9, microlotes (bajo aceite)10,11, difusión de interfaz libre (dispositivos microfluídicos)12 y diálisis (uso de botones y otras técnicas)13,14,15. Sin embargo, también existen otras configuraciones más especializadas, como los enfoques de mesofase para cristalizar proteínas de membrana16,17. Si bien la mayoría de las estructuras de proteínas de rayos X depositadas en el Banco de Datos de Proteínas se han resuelto hasta ahora mediante métodos de cristalización por difusión de vapor 6,18, otros enfoques, como la cristalización por diálisis, parecen estar infrautilizados, probablemente debido a los aspectos prácticos relacionados con su configuración experimental.
La cristalización por diálisis simplemente se basa en la lenta difusión de solutos (precipitantes, iones, aditivos y tampones) a través de una membrana semipermeable que simultáneamente impide que circulen las moléculas de proteínas. De esta manera, la solución proteica se equilibra lentamente, y el precipitante alcanza la concentración necesaria para cristalizar. La cinética del sistema depende de la temperatura, la concentración de precipitante y el corte de peso molecular de la membrana de celulosa (MWCO)19. Hasta la fecha, la configuración de cristalización más popular por diálisis ha sido el uso de botones de microdiálisis hechos de láminas de acrílico transparente. Estos generalmente se sumergen en depósitos (principalmente utilizando placas de gota colgantes de difusión de vapor) que contienen las soluciones precipitantes de cristalización. Sin embargo, este método de menor rendimiento también requiere un ensamblaje específico para sellar la solución de proteína dentro de la membrana de diálisis colocada sobre la cámara del botón, como se ilustra en la Figura 1. Además, las burbujas de aire atrapadas entre la membrana de diálisis y la solución proteica son un problema frecuente que afecta el crecimiento de cristales. Otra limitación del método son los requisitos de la muestra, por lo que se necesitan concentraciones y volúmenes mucho más altos en comparación con los métodos de difusión de vapor, para acomodar los botones de diálisis. Por lo tanto, la cristalización mediante botones de microdiálisis se ha percibido como un método poco atractivo, especialmente para objetivos difíciles como las proteínas de membrana, cuyos rendimientos de purificación son frustrantemente bajos. Recientemente, se han desarrollado dispositivos microfluídicos para facilitar la cristalización de proteínas por diálisis15. Estos chips también han sido diseñados para tener una alta transparencia de rayos X con bajo fondo, lo que permite que los chips se utilicen para la recopilación de datos in situ a temperatura ambiente, eliminando así el inconveniente de cosechar y crioenfriar cristales. A pesar de estos avances, el enfoque sigue siendo de muy bajo rendimiento y costoso.
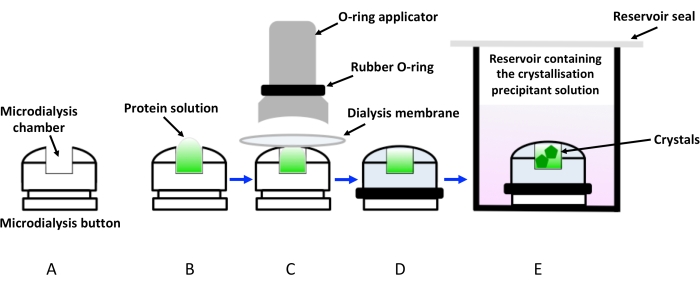
Figura 1: Representación esquemática de la cristalización por diálisis mediante botones de diálisis. (A) Representación esquemática de un botón de diálisis de cristalización. (B) La solución de proteína se agrega a la cámara de botón de microdiálisis. (C) La membrana de diálisis se sujeta al botón de microdiálisis con la ayuda de un anillo de goma (junta tórica) aplicado a través de un aplicador. (D) El botón de diálisis está listo para sumergirse en el depósito que contiene la solución de cristalización (solución de diálisis), como se muestra en (E). El vial que contiene el botón de diálisis sumergido debe sellarse para evitar la evaporación. Haga clic aquí para ver una versión más grande de esta figura.
Aquí, se presenta un protocolo sencillo para detectar las condiciones de cristalización de proteínas y el crecimiento de cristales utilizando la placa de diálisis de alto rendimiento de 96 pocillos. Estas placas desechables están diseñadas para ser utilizadas de manera similar a las placas de cristalización por difusión de vapor (pipeta y luego sello), como se muestra en la Figura 2. Las placas pueden acomodar hasta 3,2 μL de proteína y 350 μL de solución de diálisis. Cada pozo cuenta con una membrana de celulosa regenerada separada para evitar la contaminación cruzada entre los pozos. La configuración tarda alrededor de 10 minutos en completarse y no requiere ningún equipo especializado además de lo que se puede encontrar en todos los laboratorios de cristalización estándar. Se utilizan cuatro proteínas diferentes, incluidas dos proteínas de membrana, para demostrar y validar este enfoque como un método eficaz para la cristalografía de proteínas de alto rendimiento (HTP).
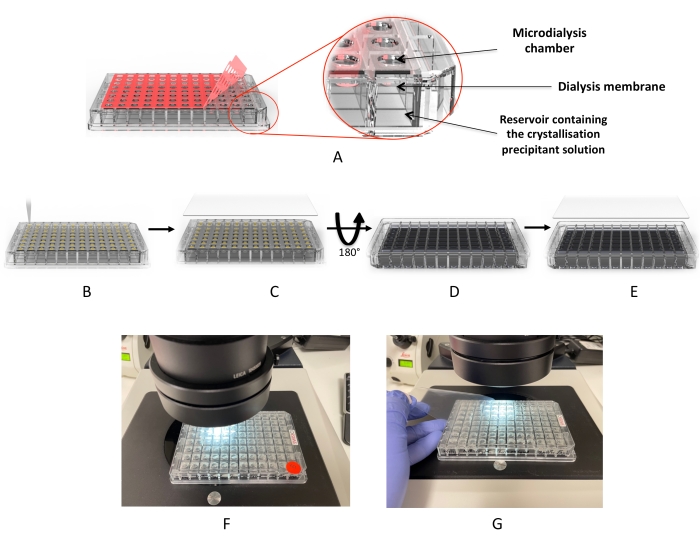
Figura 2: Flujo de trabajo de cristalización utilizando la placa de microdiálisis. (A) Eliminación de la “película de cubierta” adhesiva roja. (B) Dispensar las gotas de proteína en cada uno de los pocillos de gota. (C) Los pozos están cubiertos con la “película de cubierta” UV. (D) La placa se invierte para agregar las soluciones de diálisis (o pantalla de cristalización). (E) La placa se sella e incuba. (F,G) Inspección microscópica de las gotas. Haga clic aquí para ver una versión más grande de esta figura.
El uso de este protocolo de cristalización por diálisis se demostró utilizando el tubo dializador de 0,5 ml (Figura 3) para la producción a gran escala (cientos a miles) de microcristales, adecuados para métodos de recolección de datos de última generación, como la cristalografía en serie en ambas instalaciones XFEL 20,21,22,23,24 y sincrotrones25,26,27 , así como para los enfoques MicroED28,29,30.
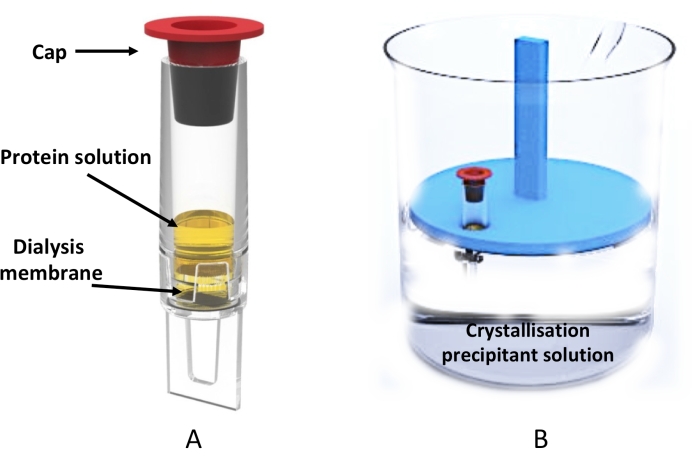
Figura 3: Cristalización de microdiálisis a gran escala utilizando el tubo dializador . (A) Representación esquemática del tubo dializador de 0,5 ml. (B) Vista lateral de un vaso de precipitados que contiene la solución de cristalización y la rejilla de tubos flotantes que sostiene un tubo de dializador. Haga clic aquí para ver una versión más grande de esta figura.
