الكائنات التمثيل الضوئي مثل الدياتومات ويجب التعامل مع ظروف الإضاءة المتغيرة والاستجابة مع آليات التأقلم متطورة للحفاظ على كفاءة التمثيل الضوئي عالية وحماية من التلف صور الأكسدة بالضوء المفرط. هو عملية كبيرة واقية من الضوء في التمثيل الضوئي حقيقيات النوى الطاقة عالية التبريد (ه) يمتص الضوء الذي يحدث كمساهمة رئيسية غير الضوئية التبريد (نبق) تحت ظروف الإجهاد الخفيفة1،2 ،3. مجمعات الهوائي الحصاد الخفيفة (LHC) تشارك في تنظيم مسارات نقل الطاقة الإثارة. في استجابة للضوء العالي الناجم عن انخفاض درجة الحموضة في التجويف بلاستيدات الخضراء، مفاتيح نظام الهوائي من ضوء حصاد الدولة إلى الدولة التبريد. وتحمي هذه الدولة المسرف الطاقة فوتوسيستيمس (PS) والمجمعات الأخرى في غشاء ثايلاكويد من التأكسد. في حقيقيات النوى التمثيل الضوئي، فعله عادة اثنين العوامل1،،من23. عامل واحد هو الضوء المتخصصة حصاد البروتين الذي يستجيب لانخفاض درجة الحموضة. يستحث البروتين الهيئاته في أعلى النباتات4. لهكسرس5، عن طريق نشاط الهيئات الفرعية الرئيسية، والتضمين الحث سه في الطحالب الخضراء6. الدياتومات تمتلك لك مثل البروتينات التي هيكلياً المتصلة لهكسرس7،،من89،10.
والعامل الثاني من سه هو دورة كزانتوفيل حيث تحويلها إلى نموذج واقية من الصور بواسطة إبوكسدة دي الكاروتينات الهوائي وعادت قبل إبوكسدة. وفي النباتات والطحالب الخضراء، يتم تحويل فيولاكسانثين إلى تين وزياكسانثين. في الدياتومات، يتم تحويل ديادينوكسانثين إلى دياتوكسانثين، الذي ثم يرتبط بمدى نبق11. ضوء دياتوم حصاد هوائي تمتلك بعض الخصائص المميزة على الرغم من أنها التطورية المتعلقة بالنباتات والطحالب لهكس. التبديل من الضوء الحصاد لحماية الصور بسرعة هائلة وقدرة نبق أعلى بالمقارنة بالنباتات12. قد يكون هذا أحد الأسباب لماذا الدياتومات ناجحة جداً في مختلف المنافذ الإيكولوجية في طريقة التي هم مسؤولون عنها ليصل إلى 45 في المائة من المحيطات صافي الإنتاج الأولى13. ومن ثم، دياتوم الخفيفة نظم الحصاد كائن مثيرة لاهتمام لبحوث عملية التمثيل الضوئي.
الدياتومات، مثل الأنواع التي تركز على مينيغينيانا سيكلوتيلا، تمتلك ثايلاكويد الخفيفة الأصيلة سميت الأصباغ أنها نظم الحصاد ربط-فوكوكسانثين، الكلوروفيل (شيلي) وج، ومن ثم الضوء FCP. حصاد البروتينات، مثل فيرفااس، هي جزءا لا يتجزأ من نظام غشاء ثايلاكويد تتألف من عدة طبقات الغشاء. الدياتومات تشكيل عصابات من ثلاث ثيلاكويدس. هذا المجمع الوضع يجعل من الصعب على دراستها على المستوى الجزيئي في غشاء ثايلاكويد. وبالإضافة إلى ذلك، العديد من العناصر المساهمة في تنظيم الضوء الحصاد (انظر أعلاه). ولذلك، في العديد من النهج، كانت المجمعات المعزولة من الغشاء باستخدام المنظفات معتدل، مثل ن-دوديسيل-β-د-مالتوبيرانوسيدي (β-DDM)، الذي جعل الغشاء ولكن الاحتفاظ مجمعات FCP سليمة. أجريت العديد من الدراسات الطيفية باستخدام solubilized FCP للتحقيق في الطاقة إينتراموليكولار نقل14،15،،من1617. غير أن هذا النهج السابق كان محدودا نظراً لتنظيم نقل الطاقة يحتاج تفاعل اكسسيتونيك مع مجمعات الهوائي أو فوتوسيستيمس أخرى. ومن ثم فهذه الأنواع من الدراسات لا تنفذ مع مجمعات solubilized نظراً للتفاعل بين المجمعات يتم فقدان.
هو سمة هامة في التنظيم هوائي “الزحام الجزيئي” هوائي وفوتوسيستيمس بغشاء ثايلاكويد18. التي أجريت سابقا، نهج بسيط لمحاكاة هذا التأثير في المختبر. تمت إزالة مواد التنظيف، مما يؤدي إلى تجميع عشوائي لمجمعات الهوائي. على الرغم من أن تم الحصول على بعض البيانات معقولة بهذا النهج،من1719، إزالة المنظفات لا تعكس الحالة في فيفو ولديها بعض القيود نظراً للمجمعات لا تتفاعل في تلك المرحلة الجامعية العادية الهيكل.
استخدام الدهنية ويتغلب على العديد من القيود السابقة. بنية التعليم العالي لا تزال سليمة تماما. غشاء الحويصلية يوفر بيئة شبه أصلية لمجمعات الهوائي. الغشاء الذي يفصل داخل الحويصلية البيئة الخارجية. بهذه الوسائل، توفر الدهنية المقصورات رد فعل اثنين للدراسات من التدرجات أيون ودرجة الحموضة، وكذلك فيما يتعلق بعمليات النقل. علاوة على ذلك، يمكن التحكم معلمات النظام التجريبي أكثر سهولة لدراسات في غشاء ثايلاكويد. وعرضت الدهنية أصلاً لتكون أداة ممتازة لدراسة المجمعات التمثيل الضوئي. وكان محورا رئيسيا في الماضي على مصنع LHC حيث تم اختبار تأثير الدهن غيرت تركيبة على الثاني LHC20. في نهج أخرى، كانت البروتين-بروتين التفاعل بين مختلف LHC ثانيا التحقيق21. أيضا، بعض الدراسات في الطحالب الخضراء نفذت تصف تجميع عفوية بين LHC22. وبالنظر إلى أهمية الدياتومات للنظم الإيكولوجية المائية، أجريت دراسات قليلة نسبيا مع مجمعات الهوائي من الدياتومات. دراستان التحقيق مجمعات الهوائي لمركزية مينيغينيانا سيكلوتيلا، وعرضت فيها تجميع الهوائي FCP23 والاستجابة من FCP للتدرجات الكهروكيميائية24 . وهكذا، الدهنية أداة ممتازة لدراسة الهوائيات دياتوم والتفاعل والتنظيم في الشروط الأصلية تقريبا. الدهنية تنوعاً منذ العديد من الشروط مثل تكوين الدهن والحويصليه حجم وكثافة البروتين ويمكن التحكم في المرحلة المائية المحيطة بها. وعلاوة على ذلك، يتطلب الأسلوب كميات قليلة من العينات. النظام التجريبي أقوى واستنساخه بدرجة عالية. تجزئة الدهنية يسمح لدراسة درجة الحموضة وأيون التدرجات، التي تتسم بأهمية العوامل في تنظيم مجمعات الهوائي.
وهنا يصف لنا عزل مجمعات الهوائي FCP من مينيغينيانا جيم وإدماجها في الدهنية مع تكوين الدهن ثايلاكويد الطبيعية. أيضا، ونحن تقديم بيانات نموذجية لتوصيف solubilized FCP الطيفية ومقارنتها مع FCP في الدهنية. ويلخص الأسلوب المعرفة والبروتوكولات الموحدة التي تم الحصول عليها من التحسينات التي غوندرومان و 2012 بوشل23وناتالي et al. عام 201622، و أحمد و 2017 ديتزيل24.
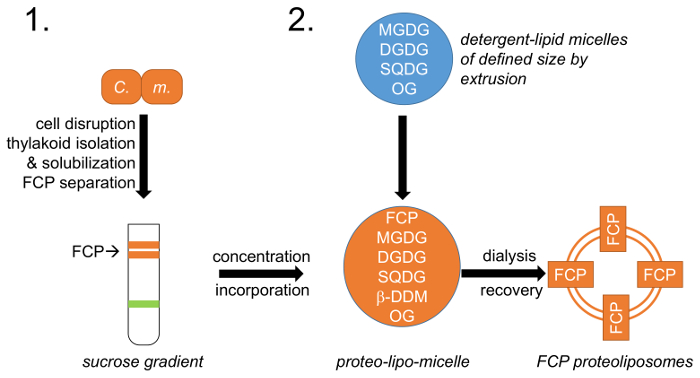
رقم 1: التمثيل التخطيطي لسير العمل. (1) يشير إلى الفقرة 1 التي تصف نمو الخلايا، واضطراب والعزلة ثايلاكويد مع عقب الانفصال FCP على السكروز كثافة التدرجات؛ جيم-م. –مينيغينيانا سيكلوتيلا الخلايا. (2) إعداد خليط دهن ثايلاكويد الطبيعية (مجدج ودجدج وسقدج) الوارد وصفها في الفقرة 2، وإنشاء المذيلات المنظفات الدهنية مع أوكتيلجليكوسيدي (الجريدة الرسمية). حجم مذيل دهن محددة يتحقق عن طريق البثق استخدام الأغشية لقطر المسام محددة. FCP والدهن-المذيلات هي موحدة في دهن مسبقاً: نسبة البروتين والمنظفات OG و β-DDM يتم إزالتها عن طريق التحكم في الغسيل الكلوي تشكيل بروتيوليبوسوميس FCP. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
