Organismos fotossintéticos como diatomáceas devem lidar com a constante mudança de condições de luz e responder com mecanismos sofisticados de aclimatação que sustentar alta eficiência fotossintética e protegem contra danos foto-oxidativo causado pela luz excessiva. Um processo de luz-protetores principal em eucariontes fotossintéticos é a alta energia têmpera (qE) da luz absorvida que ocorre como a principal contribuição para a não-fotoquímica têmpera (NPQ) sob condições de estresse leve1,2 ,3. Os complexos de antena colheita luz (LHC) estão envolvidos na regulação das vias de transferência de energia de excitação. Em resposta à luz alta induzidas pelo baixo pH no lúmen do cloroplasto, os interruptores do sistema de antena da luz colheita de estado para o estado de têmpera. Este estado de energia dissipativas protege fotossistemas (PS) e outros complexos na membrana tilacoides de foto-oxidação. Em eucariontes fotossintéticos, o qE é geralmente induzida por dois fatores1,2,3. Um fator é a luz especializada colheita proteína que responde ao baixo pH. A proteína PsbS induz o qE , em maior de plantas4. LhcSRs5, modulada pela atividade do PsbS, induzir o qE nas algas verdes6. Diatomáceas possuem proteínas, como Lhcx que estruturalmente relacionada com a LHCSRs7,8,9,10.
O segundo fator de qE é o ciclo da xantofila onde carotenoides da antena são convertidos em uma forma de foto-protetora por de epoxidação e revertidos por epoxidação. Plantas e algas verdes, violaxantina é convertida em zeaxantina. Diatomáceas, diadinoxantina é convertida em diatoxanthin, que em seguida se correlaciona com a extensão da NPQ11. A luz de diatomáceas colheita antena possui algumas peculiaridades, embora relacionadas com plantas e algas LHCs é evolutiva. O interruptor de luz da colheita para foto-proteção é extremamente rápido e a capacidade NPQ é superior em comparação com plantas12. Isto pode ser uma razão porque diatomáceas são muito bem sucedidas em diferentes nichos ecológicos, de forma que eles são responsáveis por até 45% da produção primária líquida oceânica13. Portanto, luz, sistemas de colheita de diatomáceas são um interessante objeto de pesquisa de fotossíntese.
Diatomáceas, como as espécies centralizado no Cyclotella meneghiniana, possuem tilacoides intrínseca luz colheita sistemas em homenagem os pigmentos eles vincular – fucoxantina, clorofila (chl) a e c, daí Light FCP. colheita de proteínas, tais como FCPs, são incorporado no sistema de membrana tilacoides, composto por várias camadas de membrana. Diatomáceas formam bandas de três contém. Este complexo situação dificulta a estudá-los no nível molecular na membrana tilacoides. Além disso, muitos componentes contribuem para a regulação da luz da colheita (veja acima). Portanto, em muitas abordagens, os complexos foram isolados a partir da membrana usando detergentes suaves, tais como n-dodecil-β-D-maltopyranoside (β-DDM), que solubilizar a membrana, mas manter os complexos FCP intacta. Muitos estudos espectroscópicos foram realizados usando o FCP solubilizado para investigar a energia intramolecular transferência14,15,16,17. No entanto, esta primeira abordagem foi limitada, desde que o Regulamento de transferência de energia precisa de excitônicas interação com outros complexos de antena ou de fotossistemas. Daí, esses tipos de estudos não podem ser realizados com solubilizado complexos porque a interação entre complexos é perdida.
Uma característica importante no Regulamento da antena é a “exclusão molecular” da antena e fotossistemas na membrana tilacoides18. Anteriormente, realizou uma abordagem simples para simular este efeito in vitro. O detergente foi removido, o que leva à agregação aleatória de complexos de antena. Embora alguns dados razoáveis foi obtidos por esta abordagem17,19, a remoção do detergente não reflete a situação na vivo e tem algumas limitações, desde que os complexos não estão interagindo em sua regular terciário estrutura.
O uso de lipossomas supera várias das limitações anteriores. A estrutura terciária ainda está totalmente intacta. A membrana do lipossoma fornece um ambiente quase nativo para os complexos de antena. A membrana separa o interior do lipossoma o ambiente exterior. Por estes meios, os lipossomas fornecem dois compartimentos de reação para estudos de gradientes iônica e pH, bem como quanto aos processos de transporte. Além disso, os parâmetros do sistema experimental podem ser controlados mais facilmente para estudos na membrana tilacoides. Os lipossomas foram já mostrou ser uma excelente ferramenta para o estudo de complexos fotossintéticos. Um grande foco no passado foi na planta LHC onde o efeito de composição de lipídios alterados foi testado no LHC II20. Em outras abordagens, a interação da proteína-proteína entre diferentes LHC II foram investigados21. Além disso, alguns em algas verdes foram realizados estudos que descrevem a aglomeração espontânea entre LHC22. Considerando a importância das diatomáceas para ecossistemas aquáticos, relativamente poucos estudos foram realizados com complexos de antena de diatomáceas. Dois estudos investigaram os complexos antena do centralizado no meneghiniana de Cyclotella, onde a aglomeração do FCP antena23 e capacidade de resposta do FCP para gradientes eletroquímicos24 foram mostrados. Assim, os lipossomas são uma excelente ferramenta para estudar a antenas de diatomáceas e sua interação e regulamento em condições quase nativos. Os lipossomas são versáteis desde muitas condições tais como a composição de lipídios, lipossomas tamanho, densidade de proteína e a fase aquosa circundante pode ser controlada. Além disso, o método requer pequenas quantidades de amostras. O sistema experimental é robusto e altamente reprodutíveis. Compartimentação dos lipossomas permite estudar o pH e gradientes de íons, que são importantes fatores na regulação dos complexos de antena.
Aqui, descrevemos o isolamento de complexos de antena FCP de c. meneghiniana e sua incorporação em lipossomas com composição lipídica de tilacoides natural. Também, fornecemos dados exemplares para a caracterização espectroscópica de FCP solubilizado e compará-los com FCP em lipossomas. O método resume o conhecimento e protocolos padronizados obtidos com as melhorias de Gundermann e Büchel 201223Natali et al 201622e Ahmad e 2017 Dietzel24.
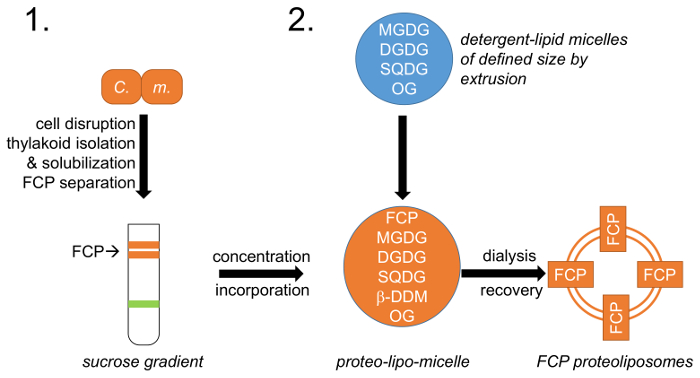
Figura 1: representação esquemática do fluxo de trabalho. (1) refere-se ao n º 1, que descreve o crescimento celular, ruptura e isolamento de tilacoides com a seguinte separação FCP em gradientes de densidade de sacarose; C. m. -Células deCyclotella meneghiniana . (2) preparação de tilacoides natural mistura de lipídios (MGDG, Junior e SQDG) descrita no n º 2 e criação de micelas de lipídios-detergente com octylglycoside (OG). Um tamanho definido lipid-micelle é alcançado por extrusão usando membranas de um diâmetro de poro definido. FCP e micelas de lipídios são unificadas em um lipido predefinido: proporção de proteína e os detergentes OG e β-DDM são removidos através de controlada diálise formando FCP proteoliposomes. Clique aqui para ver uma versão maior desta figura.
