Fotosentetik canlılar Diyatomlarla gibi sürekli değişen ışık koşulları ile başa çıkmak ve yüksek fotosentetik verimliliği sürdürmek ve aşırı ışık tarafından fotoğraf-oksidatif hasarı korumak sofistike calıştıkları mekanizmaları ile yanıt. Bir büyük ışık-koruyucu fotosentetik ökaryotlarda yüksek enerji su verme (qE) olmayan fotokimyasal su verme (NPQ) hafif stres koşulları1,2 altında ana katkısı olarak oluşur emilen ışık işlemidir ,3. Işık hasat Anten kompleksleri (LHC) uyarma enerji transferi yolları Yönetmelikte katılmaktadırlar. Yanıt yüksek ışık olarak düşük pH kloroplast lümen, anten sistem anahtarları devlet su verme durumuna hasat ışık indüklenen. Bu enerji dissipative devlet photosystems (PS) ve diğer kompleksleri thylakoid zarda fotoğraf-oksidasyon korur. Fotosentetik ökaryotlarda qE genellikle iki faktör1,2,3tarafından indüklenen. Düşük pHiçin yanıt protein hasat özel ışık bir faktördür. PsbS protein yüksek bitkiler4qE neden olmaktadır. LhcSRs5PsbS faaliyete göre modüle, neden yeşil algler6‘E q. Diyatomlarla yapısal olarak LHCSRs7,8,9,10‘ a ilgili Lhcx gibi proteinler sahip.
İkinci qE nerede karotenoidler anten fotoğraf-koruyucu forma de-epoxidation tarafından dönüştürülür ve epoxidation tarafından yeniden dönmek xanthophyll döngüsü faktördür. Bitki ve yeşil algler, violaxanthin zeaksantin için dönüştürülür. Diyatomlarla, diadinoxanthin diatoxanthin, sonra NPQ11ölçüde ile karşılıklı olarak ilişkilendirir dönüştürülür. Bitki ve yosun LHCs ilgili evrimsel olsa anten hasat diatom ışık bazı özelliklerine sahiptir. Fotoğraf-koruma için hasat ışık geçiş son derece hızlı ve NPQ kapasitesi daha bitkiler12‘ ye göre. Bu neden Diyatomlarla okyanus net birincil üretim13ilâ % 45 sorumlu oldukları bir şekilde farklı ekolojik niş içinde çok başarılı bir nedeni olabilir. Bu nedenle, diatom sistemleri hasat ışık fotosentez araştırma ilginç bir nesne vardır.
Merkezli türler Cyclotella meneghiniana, thylakoid içsel ışık sistemleri sonra pigmentler adlı hasat sahip gibi Diyatomlarla – fucoxanthin, klorofil (chl) a ve c, dolayısıyla FCP proteinler, hasat ışık bağlama FCPs gibi vardır birçok membran katmandan oluşan thylakoid membran sisteminde gömülü. Diyatomlarla üç thylakoids grupları oluşturur. Bu karmaşık durum thylakoid zarda moleküler düzeyde çalışmaya zor yapar. Buna ek olarak, birçok bileşenleri (yukarı bakın) hasat ışık düzenlenmesi için katkıda bulunur. Bu nedenle, pek çok yaklaşım içinde kompleksleri gibi n-Lauryl-β-D-membran solubilize ama FCP kompleksleri sağlam tutmak maltopyranoside (β-demir), hafif deterjan kullanarak zar izole edildi. İntramolecular enerji transferi14,15,16,17araştırmak için çözündürüldükten FCP kullanarak birçok spektroskopik çalışmalar gerçekleştirilmiştir. Enerji transferi Yönetmeliği ile diğer Anten kompleksleri veya photosystems excitonic etkileşim ihtiyacı ancak, bu eski yaklaşım sınırlı alınmaya başlanmıştır. Kompleksleri arasındaki etkileşimi kayıp olduğu için bu nedenle, bu tür çalışmalar çözündürüldükten kompleksleri ile uygulanmaz.
Önemli bir anten Yönetmelikte “moleküler kalabalık” anten ve thylakoid membran18photosystems özelliğidir. Eskiden, basit bir yaklaşım bu etkisi simüle etmek için yapılmıştır tüp bebek. Anten kompleksleri rasgele toplama için bu da neden deterjan kaldırıldı. Her ne kadar bazı makul veri bu yaklaşım17,19tarafından elde edildi, deterjan kaldırma durumu vivo içinde yansıtmaz ve kompleksleri onların düzenli Tersiyer etkileşim değil bu yana bazı sınırlamalar vardır yapısı.
Lipozomlar kullanımı birkaç eski sınırlamalar üstesinden gelir. Üçüncül yapı hala tam olarak sağlamdır. Lipozom membran Anten kompleksleri için yarı doğal bir ortam sağlar. Membran lipozom iç dış ortamdan ayıran. Bu şekilde, lipozomlar iyon ve pH degradeleri de taşıma işlemleri gelince, çalışmalar için iki reaksiyon bölmeler sağlar. Ayrıca, deneysel sistem parametrelerini daha kolay thylakoid membran araştırmaları kontrol edilebilir. Lipozomlar zaten fotosentetik kompleksleri eğitim için mükemmel bir araç olarak gösterildi. Geçmişte büyük bir odak bitki nerede değişmiş lipid kompozisyon etkisini LHC II20üzerinde test edildi LHC oldu. Diğer yaklaşımlar, farklı LHC II arasındaki protein-protein etkileşimi incelenen21idi. Ayrıca, yeşil algler bazı çalışmalarda spontan LHC22arasında kümeleme açıklayan gerçekleştirilmiştir. Diyatomlarla sucul ekosistemler için önemi göz önüne alındığında, görece az sayıda çalışmalar Diyatomlarla Anten kompleksleri ile gerçekleştirilmiştir. İki çalışma nerede FCP anten23 ve FCP yanıt elektrokimyasal gradyan24 için kümeleme gösterildi merkezli Cyclotella meneghiniana, Anten kompleksleri araştırıldı. Böylece, lipozomlar diatom antenler ve etkileşim ve neredeyse doğal koşullar Yönetmelikte eğitim için mükemmel bir araç vardır. Lipozomlar lipid bileşimi, lipozom boyutu, protein yoğunluğu gibi birçok koşulları beri çok yönlü ve çevresindeki sulu faz kontrol edilebilir. Ayrıca, yöntem örnekleri düşük miktarda gerektirir. Deneysel sistem sağlam ve son derece tekrarlanabilir. PH eğitimi için lipozomlar compartmentalization sağlar ve önemli iyon degradeler faktörler Anten kompleksleri Yönetmelikte.
Burada, FCP Anten kompleksleri C. meneghiniana üzerinden yalıtım ve onların birleşme lipozomlar doğal thylakoid lipid bileşimi ile açıklanmaktadır. Ayrıca, çözündürüldükten FCP spektroskopik karakterizasyonu için örnek veri sağlamak ve onları lipozomlar FCP ile karşılaştırın. Yöntem bilgi ve Gundermann ve Büchel 201223, Natali vd. 201622ve Ahmed ve Dietzel 201724ilerleme–dan elde edilen standart iletişim kuralları özetler.
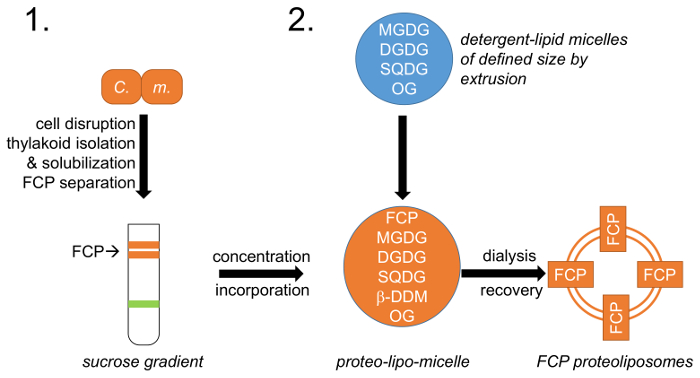
Şekil 1: iş akışı şematik gösterimi. (1) başvuran hücre büyüme, bozulma ve FCP ayrılık sükroz yoğunluk gradyanlar üzerinde; aşağıdaki thylakoid yalıtım tanımlayan paragraf 1 C. m. –Cyclotella meneghiniana hücreleri. (2) doğal thylakoid lipid karışımı (MGDG, DGDG ve SQDG) hazırlanması 2 ve lipid-deterjan micelles octylglycoside (OG) ile oluşturulması paragrafta açıklanan. Tanımlanmış lipid-micelle boyutu ekstrüzyon tarafından tanımlanmış gözenek çapı membranlar kullanılarak elde edilir. FCP ve lipid-micelles önceden tanımlanmış bir lipid birleştirilmiş: protein oranı ve OG ve β-DDM deterjanlar vardır FCP proteoliposomes oluşturan kontrollü kaldırılan yolu ile diyaliz. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
