Organismos fotosintéticos tales como diatomeas deben lidiar con condiciones de luz cambiantes y responder con mecanismos sofisticados de aclimatación que mantener alta eficiencia fotosintética y protegen de daño foto-oxidativo causado por la luz excesiva. Un importante proceso de luz protectora en eucariotas fotosintéticos es la alta energía amortiguamiento (qE) de la luz absorbida que se presenta como la principal contribución a la Temple no fotoquímica (NPQ) bajo condiciones de estrés ligero1,2 ,3. Los complejos antena cosecha luz (LHC) están implicados en la regulación de vías de transferencia de energía de excitación. En respuesta a la luz alta inducida por el bajo pH en el lumen del cloroplasto, los interruptores del sistema de antena de la cosecha de estado al estado apagando la luz. Este estado disipativo de energía protege fotosistemas (PS) y otros complejos en la membrana del thylakoid de la foto-oxidación. En eucariotas fotosintéticos, qE es generalmente inducida por dos factores1,2,3. Un factor es la luz especializada recolección proteína que responde a bajo pH. La proteína subsidiarios induce la qE en las plantas superiores4. LhcSRs5, modulado por la actividad de subsidiarios, inducir la qE en algas verdes6. Las diatomeas poseen Lhcx-como proteínas relacionadas estructuralmente con LHCSRs7,8,9,10.
El segundo factor de qE es el ciclo de la xantofila donde los carotenoides de la antena son convertidos en forma de foto protección de-epoxidación y revertidos por epoxidación. En plantas y algas verdes, violaxantina se convierte en zeaxantina. En diatomeas, diadinoxanthin se convierte en diatoxanthin, que luego se correlaciona con el grado de NPQ11. La luz de la diatomea cosecha antena posee algunas peculiaridades aunque es evolutivo relacionado con plantas y algas LHCs. El interruptor de la luz cosecha a fotoprotección es enormemente rápido y la capacidad NPQ es superior comparada con las plantas12. Esta podría ser una razón por la cual las diatomeas muy exitosas en diferentes nichos ecológicos de manera que son responsables de hasta un 45% de la producción primaria neta oceánico13. Por lo tanto, luz cosecha sistemas de diatomeas son un interesante objeto de investigación de la fotosíntesis.
Diatomeas, como las centradas en especies de Cyclotella meneghiniana, poseen tilacoides luz intrínseca cosecha sistemas nombrados después de los pigmentos unen – fucoxantina, clorofila (chl) a y c, por lo tanto luz FCP. recolección de proteínas, como la FCPs, son incrustado en el sistema de membranas de tilacoides que comprende varias capas de membrana. Las diatomeas forman bandas de tres tilacoides. Este complejo situación dificulta estudiar en el nivel molecular en la membrana de los tilacoides. Además, muchos componentes contribuyen a la regulación de la luz cosecha (véase arriba). Por lo tanto, en muchos enfoques, los complejos fueron aislados de la membrana usando detergentes suaves, tales como n-dodecil-β-D-maltopyranoside (β-DDM), que solubilizan la membrana pero mantener intacto los complejos FCP. Se realizaron muchos estudios espectroscópicos uso FCP solubilizado para investigar energía intramolecular transferencia14,15,16,17. Sin embargo, este enfoque anterior era limitada ya que la regulación de la transferencia de energía necesita interacción excitonic con otros complejos de antena o fotosistemas. Por lo tanto, este tipo de estudios no se puede realizar con complejos solubles debido a la interacción entre complejos se pierde.
Una característica importante en la regulación de la antena es la “apretadura molecular” de la antena y fotosistemas en la membrana tilacoides del18. Anteriormente, se llevó a cabo un enfoque simple para simular este efecto en vitro. El detergente fue quitado, que conduce a la agregación aleatoria de complejos antena. Aunque se obtuvieron algunos datos razonables por este enfoque17,19, detergente no refleja la situación en vivo y tiene algunas limitaciones ya que los complejos no están interactuando en los terciarios regulares estructura.
El uso de liposomas supera varias de las limitaciones anteriores. La estructura terciaria sigue siendo totalmente intacta. La membrana del liposoma proporciona un entorno cuasi-nativo para los complejos de antena. La membrana separa el interior de la liposoma del ambiente exterior. Por estos medios, liposomas proporcionan dos compartimientos de la reacción para estudios de gradientes de iones y pH, así como por procesos de transporte. Además, pueden controlarse más fácilmente los parámetros del sistema experimental de estudios de la membrana del thylakoid. Liposomas ya fueron demostrados para ser una excelente herramienta para el estudio de complejos fotosintéticos. Un enfoque principal en el pasado fue en la planta donde se probó el efecto de la composición lipídica alterada en LHC II20LHC. En otros enfoques, interacción de la proteína-proteína entre diferentes LHC II fueron investigados21. También, algunos en algas verdes se realizaron estudios que describen la agrupación espontánea entre el LHC22. Considerando la importancia de las diatomeas para los ecosistemas acuáticos, relativamente pocos estudios se realizaron con los complejos antena de diatomeas. Dos estudios investigaron los complejos antena de la centrada en la Cyclotella meneghiniana, donde se muestra el agrupamiento de la FCP antena23 y capacidad de respuesta de FCP a gradientes electroquímicos24 . Así, los liposomas son una excelente herramienta para el estudio de diatomeas antenas y su interacción y regulación en condiciones casi nativas. Los liposomas son versátiles desde muchas condiciones tales como la composición de lípido, liposomas tamaño, densidad de la proteína y la fase acuosa circundante puede ser controlada. Además, el método requiere pequeñas cantidades de muestras. El sistema experimental es robusto y altamente reproducibles. Permite la compartimentalización de liposomas para el estudio de pH y los gradientes de iones, que son importantes factores en la regulación de los complejos antena.
Aquí, describimos el aislamiento de complejos de antena FCP de C. meneghiniana y su incorporación en los liposomas con composición de lípido natural tilacoides. También, proporcionamos datos ejemplares para la caracterización espectroscópica de FCP solubilizado y compararlos con FCP en liposomas. El método resume conocimientos y protocolos estandarizados de las mejoras de Gundermann y Büchel 201223, Natali et al 201622y Ahmad y Dietzel 201724.
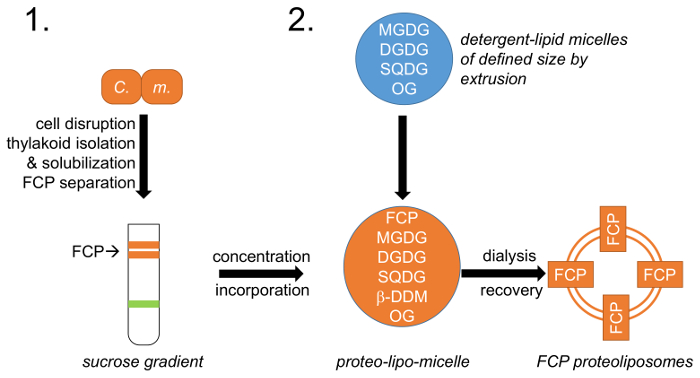
Figura 1: representación esquemática del flujo de trabajo. (1) se refiere al párrafo 1 que describe el crecimiento celular, desorganización y aislamiento de tilacoides con la siguiente separación de FCP en gradientes de densidad de sacarosa; M. C. -Células deCyclotella meneghiniana . (2) preparación de la mezcla de lípidos naturales tilacoides (MGDG, DGDG y SQDG) había descrito en el párrafo 2 y creación de micelas de detergente de lípidos con octylglycoside (OG). Se consigue un tamaño de micela de lípidos definidos por extrusión utilizando membranas de diámetro de poro definido. FCP y lípido micelas se unifican en un lípido predefinido: relación y los detergentes OG y β-DDM son quitado vía controlado diálisis formando proteoliposomes FCP. Haga clic aquí para ver una versión más grande de esta figura.
