היכולת מהנדס הגנום של כל אורגניזם חי יש לא מעט אפליקציות ביו, הביו-טכנולוגיה, כגון התיקון של גורמי מחלות מוטציות, בניית דגמי הסלולר מדויק לחקר המחלה, או דור של החקלאי יבולים עם תכונות רצויות. מאז המאה ה, טכנולוגיות שונות פותחו עבור גנום הנדסה בתאי יונקים, כולל meganucleases1,2,3, אבץ אצבע nucleases4,5, או תמלול אפקטור activator דמוי nucleases (TALENs)6,7,8,9. עם זאת, טכנולוגיות קודמות אלה הם קשה לתוכנית או מייגע להרכיב, ובכך פוגעות שלהם אימוץ נרחב מחקר ובתעשייה.
בשנים האחרונות, באופן קבוע באשכולות interspaced חזרה palindromic קצר (CRISPR) – מערכת CRISPR-הקשורים (Ca) התפתחה גנום חדש חזק הנדסה טכנולוגיה10,11. במקור מערכת חיסונית אדפטיבית בחיידקים, עבר בהצלחה לפריסה הגנום שינוי בצמחים ובעלי חיים, כולל האדם. הסיבות העיקריות מדוע CRISPR-Cas צברה פופולריות רבה בזמן כל כך קצר זה הרכיב המביאה את endonuclease Cas מפתח, כגון Cas9 או Cas12a (ידוע גם בשם Cpf1), על המיקום הנכון הגנום הוא פשוט חתיכת קצר מדריך בודד chimeric RN (SgRNA), אשר היא פעולה פשוטה עיצוב וזול לסנתז. לאחר לאתר היעד האנזים Cas מתפקד בתור זוג מספריים מולקולרית ודבק ה-DNA מאוגד עם שלו RuvC, HNH או Nuc תחומים12,13,14. מעבר נטושים כפול וכתוצאה מכך (DSB) לאחר מכן תיקון על ידי התאים באמצעות קצה שאינם הומולוגיים להצטרף (NHEJ) או מסלול תיקון מכוון הומולוגיה (HDR). בהיעדרו של תבנית תיקון, DSB יתוקן על ידי השביל NHEJ לשגיאות, אשר יכולים להצמיח אקראיים מדומים ההוספה או המחיקה של נוקלאוטידים (indels) באתר לחתוך, ובכך לגרום frameshift מוטציות בגנים חלבונים. עם זאת, בנוכחות התורם תבנית המכילה את השינויים הרצויים של הדנ א, DSB יתוקן על ידי השביל HDR דיוק גבוה. סוגים נפוצים של תבניות התורם כוללים חד גדילי oligonucleotides (ssODNs), פלסמידים. לשעבר משמש בדרך כלל אם השינויים DNA המיועד קטנות (לדוגמה, שינוי של בסיס יחיד זוג רשימות), בזמן האחרון זה משמש בדרך כלל אם אנחנו רוצים להוסיף רצף ארוך יחסית (לדוגמה, רצף קידוד של חלבון פלואורסצנטי ירוק או GFP) לתוך מיקומה היעד.
הפעילות endonuclease של החלבון Cas דורש הנוכחות של protospacer הסמוך מוטיב (פאם) ב אתר היעד15. פאם של Cas9 נמצא בקצה 3′ של protospacer, פאם Cas12a (הנקראת גם Cpf1) נמצאת בקצה 5′ במקום16. Cas-המדריך RNA מורכב אין אפשרות להציג DSB אם פאם הוא נעדר17. לפיכך, פאם המקומות אילוץ על המיקומים גנומית איפה מסוגל לבקע נוקלאז Cas מסוים. למרבה המזל, nucleases Cas של מינים שונים של חיידקים בדרך כלל התערוכה פאם דרישות שונות. לפיכך, על ידי שילוב של המערכות CRISPR-Cas השונות שלנו ארגז כלים הנדסיים, אפשר להרחיב את מגוון אתרים אשר עשוי להיות ממוקד הגנום. יתר על כן, אנזים Cas טבעי שניתן הנדסה לאחור או התפתחו כדי לזהות רצפים פאם חלופיים, שמפרידה את היקף נגיש מניפולציה18,19,20גנומית מטרות.
למרות מספר מערכות CRISPR-Cas זמינים למטרות הנדסה הגנום, רוב המשתמשים של הטכנולוגיה יש הסתמך בעיקר על נוקלאז Cas9 מ הפישחה ינפל תומימת סטרפטוקוקוס (SpCas9) מסיבות רבות. קודם כל, זה דורש יחסית פשוט הגולה פאם, בניגוד רבים לחלבונים אחרים-Cas זה יכול לבקע רק בנוכחות PAMs מורכבים יותר. שנית, זה endonuclease Cas הראשון כדי לפרוס בהצלחה תאים אנושיים21,22,23,24. שלישית, SpCas9 הוא האנזים מאופיין בצורה הטובה ביותר עד כה. אם חוקר מבקש להשתמש נוקלאז Cas אחר, הוא או היא לעתים קרובות יהיה ברור על הדרך הטובה ביותר לעצב את הניסוי ותבצע כמה אנזימים אחרים בהקשרים ביולוגיים שונים, בהשוואה ל- SpCas9.
כדי להוסיף הבהרות לגבי את הביצועים היחסיים של מערכות CRISPR-Cas שונות, לאחרונה ערכנו השוואה שיטתית של חמש רשויות אישורים endonucleases – SpCas9, האנזים Cas9 מ Staphylococcus aureus (SaCas9), האנזים Cas9 מ- מנינגוקוקוס (NmCas9), האנזים Cas12a מן Acidaminococcus sp. BV3L6 (AsCas12a), האנזים Cas12a של החיידק Lachnospiraceae ND2006 (LbCas12a)25. להשוואה הוגן, הערכנו את nucleases רשויות אישורים שונים באמצעות אותה קבוצה של אתרי יעד ותנאים אחרים ניסיוני. המחקר גם מאפשרת עיצוב הפרמטרים עבור כל מערכת CRISPR-Cas, אשר ישמש כהפניה שימושי עבור משתמשים של הטכנולוגיה. . הנה, טוב יותר לאפשר לחוקרים לנצל CRISPR-Cas מערכת, אנו מספקים פרוטוקול צעד אחר צעד עבור הנדסה הגנום אופטימלית עם אנזימים שונים Cas9 ו- Cas12a (ראה איור 1). הפרוטוקול כוללת לא רק פרטים ניסיוני אבל שיקולי התכנון חשוב גם כדי להגדיל את הסבירות של תוצאה הנדסה מוצלחת הגנום בתאים בתרבית.
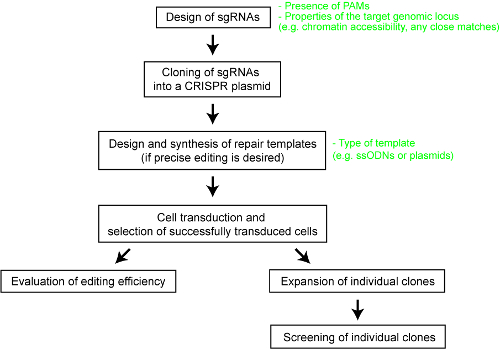
איור 1 : סקירה כללית של זרימת העבודה כדי ליצור הגנום נערך שורות תאים אנושיים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
