La capacidad para diseñar el genoma de cualquier organismo vivo tiene muchas aplicaciones biomédicas y biotecnológicas, como la corrección de enfermedad-causar mutaciones, construcción de modelos celulares precisos estudios de la enfermedad, o generación de agrícola cultivos con características deseables. Puesto que la vuelta del siglo, se han desarrollado diversas tecnologías para la ingeniería del genoma en células de mamíferos, incluyendo meganucleases1,2,3, zinc finger nucleasas4,5, o transcripción efector activador como nucleasas (TALENs)6,7,8,9. Sin embargo, estas tecnologías anteriores son programa difícil o tedioso de montar, tal modo obstaculizando su adopción generalizada en la investigación y la industria.
En los últimos años, el cluster regularmente otro corto repite palindrómico (CRISPR) – sistema CRISPR-asociado (Cas) se ha convertido en un genoma nuevo potente ingeniería tecnología10,11. Originalmente un sistema inmune adaptante en las bacterias, ha sido exitosamente implementado para la modificación del genoma en plantas y animales, incluyendo seres humanos. La razón principal por qué CRISPR-Cas ha ganado tanta popularidad en tan corto tiempo es el elemento que trae la clave endonucleasa de Cas, como Cas9 o Cas12a (también conocido como Cpf1), en la ubicación correcta en el genoma es simplemente una pequeña pieza de guía solo quimérica RN A (sgRNA), que es de diseño sencillo y barato sintetizar. Después de ser reclutado al sitio de destino, la enzima Cas funciona como un par de tijeras moleculares y hiende la DNA dependiente con su RuvC, HNH o Nuc dominios12,13,14. Posteriormente se repara la rotura trenzada doble resultante (de diferencias OSD) por las células a través de la vía de reparación dirigido por homología (HDR) o no homóloga se terminan uniendo (NHEJ). En la ausencia de una plantilla de reparación, el OSD es reparado por la vía NHEJ propenso, que puede dar lugar a pseudo-random inserción o deleción de nucleótidos (indels) en el sitio de corte, potencialmente causando mutágeno ‘ frameshift ‘ mutaciones en genes de la proteína-codificación. Sin embargo, en presencia de una plantilla de donante que contiene los cambios de ADN deseados, el OSD es reparado por la vía HDR de alta fidelidad. Tipos comunes de donantes plantillas son oligonucleótidos de cadena simple (ssODNs) y plásmidos. El primero se utiliza normalmente si los cambios previstos de DNA son pequeños (por ejemplo, alteración de un solo par de base), mientras que este último se suele utilizar si se desea insertar una secuencia relativamente larga (por ejemplo, la secuencia de codificación de una proteína verde fluorescente o GFP) en el lugar de destino.
La actividad endonucleasa de la proteína Cas requiere la presencia de un motivo adyacente de protospacer (PAM) en el sitio de destino15. PAM de Cas9 es el extremo 3′ de la protospacer, mientras que el PAM de Cas12a (también llamado Cpf1) está en el extremo 5′ en lugar16. La guía de Cas RNA complejo es incapaz de introducir un OSD si el PAM es ausente17. Por lo tanto, el PAM coloca una restricción en las localizaciones genómicas donde una nucleasa Cas particular es capaz de unirse. Afortunadamente, las nucleasas de la Cas de diferentes especies bacterianas típicamente exhiben diversos requisitos de PAM. Por lo tanto, al integrar varios sistemas de CRISPR-Cas en nuestra caja de herramientas de ingeniería, podemos ampliar la gama de sitios que pueden ser objeto de un genoma. Además, una enzima natural de la Cas puede ser diseñado o evolucionado para reconocer secuencias de PAM alternativas, ampliando aún más el alcance de objetivos genómicas accesibles a la manipulación18,19,20.
Aunque varios sistemas CRISPR-Cas están disponibles para propósitos de ingeniería del genoma, la mayoría de usuarios de la tecnología ha dependido principalmente la nucleasa Cas9 de Streptococcus pyogenes (SpCas9) por múltiples razones. En primer lugar, requiere un PAM relativamente simplemente NGG, a diferencia de muchas otras proteínas de Cas que pueden unirse sólo en presencia de PAM más compleja. En segundo lugar, es la primera endonucleasa de Cas a ser implementado con éxito en células humanas21,22,23,24. En tercer lugar, SpCas9 es la enzima mejor caracterizada hasta la fecha. Si un investigador desea utilizar otro nucleasa de Cas, él o ella a menudo sería confusa acerca de la mejor manera de diseñar el experimento y así otras enzimas llevará a cabo en diferentes contextos biológicos comparados con SpCas9.
Para dar claridad al desempeño relativo de los diferentes sistemas de CRISPR-Cas, recientemente hemos realizado una comparación sistemática de cinco endonucleases de Cas-SpCas9, la enzima Cas9 de Staphylococcus aureus (SaCas9), la enzima Cas9 de Meningitidis de la Neisseria (NmCas9), la enzima Cas12a de Acidaminococcus SP. BV3L6 (AsCas12a) y la enzima Cas12a de Lachnospiraceae bacteria ND2006 (LbCas12a)25. Para una comparación justa, se evaluaron las diversas nucleasas Cas mediante el mismo conjunto de sitios de destino y otras condiciones experimentales. Los parámetros de diseño del estudio también delineado para cada sistema CRISPR-Cas, que serviría como una referencia útil para los usuarios de la tecnología. Aquí, mejor permiten a los investigadores a hacer uso de la Cas CRISPR sistema, proporcionamos un protocolo paso a paso para la ingeniería del genoma óptima con los distintos enzimas Cas9 y Cas12a (ver figura 1). El Protocolo no sólo incluye detalles experimentales pero también importantes consideraciones para maximizar la probabilidad de un resultado de ingeniería exitosa del genoma en células de mamíferos.
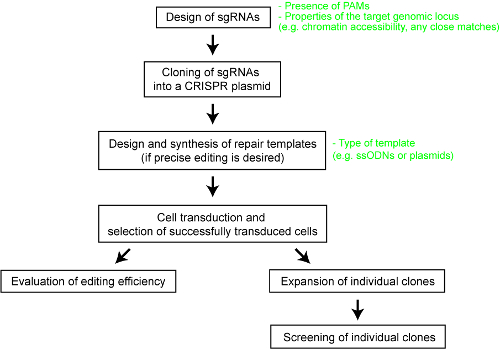
Figura 1 : Una visión general del flujo de trabajo para generar el genoma editado líneas celulares humanas. Haga clic aquí para ver una versión más grande de esta figura.
