La capacità di progettare il genoma di ogni organismo vivente ha molte applicazioni biomediche e biotecnologiche, come la correzione di malattia-causare mutazioni, costruzione di accurati modelli cellulari per lo studio di malattia, o la generazione di agricolo colture con caratteristiche desiderabili. Poiché la girata del secolo, diverse tecnologie sono state sviluppate per l’ingegneria del genoma in cellule di mammiferi, tra cui meganucleases1,2,3, zinco dito nucleasi4,5, o trascrizione dell’effettore attivatore-come nucleasi (TALENs)6,7,8,9. Tuttavia, queste tecnologie precedenti sono difficili da programma o noioso da montare, quindi che ostacolano l’adozione diffusa nella ricerca e nell’industria.
Negli ultimi anni, il cluster regolarmente intervallate brevi ripetizioni palindromi (CRISPR) – sistema di CRISPR-collegata (Cas) è emerso come un potente nuovo genoma ingegneria tecnologia10,11. Originariamente un sistema immunitario adattativo nei batteri, è stato correttamente distribuito per la modificazione del genoma in piante e animali, compreso gli esseri umani. Un motivo principale perché CRISPR-Cas ha guadagnato popolarità così tanto in così poco tempo che è l’elemento che porta la chiave dell’endonucleasi di Cas, ad esempio Cas9 o Cas12a (noto anche come Cpf1), nella posizione corretta nel genoma è semplicemente un breve pezzo di guida singola chimerico RN A (sgRNA), che è al design semplice ed economico per sintetizzare. Dopo essere reclutato nel sito di destinazione, l’enzima Cas funziona come un paio di forbici molecolari e si unirà il DNA associato con relativi RuvC, HNH o Nuc domini12,13,14. La risultante doppia interruzione incagliato (DSB) è più riparato dalle cellule via via regia di omologia repair (HDR) o non-omologo fine unirsi (NHEJ). In assenza di un modello di riparazione, il DSB è riparato dalla via NHEJ errori, che possa dar luogo a pseudo-caso inserimento o l’eliminazione di nucleotidi (indels) presso il sito di taglio, causando potenzialmente frameshift mutazioni in geni di proteina-codificazione. Tuttavia, in presenza di un modello di donatore che contiene le modifiche desiderate di DNA, il DSB è riparato dal pathway HDR ad alta fedeltà. Comuni tipi di modelli di donatore includono oligonucleotidi a singolo filamento (ssODNs) e plasmidi. Il primo viene in genere utilizzato se i previsti cambiamenti del DNA sono piccoli (per esempio, alterazione di un singolo paio di base), mentre quest’ultimo viene in genere utilizzato se si vuole inserire una sequenza relativamente lunga (ad esempio, la sequenza di codificazione di una proteina fluorescente verde o GFP) in luogo del bersaglio.
L’attività endonucleasica della proteina Cas richiede la presenza di un motivo di protospacer adiacenti (PAM) presso il sito di destinazione15. Il PAM di Cas9 è all’estremità 3′ del protospacer, mentre il PAM di Cas12a (chiamato anche Cpf1) è invece all’estremità 5′16. La Cas-guida RNA complesso è in grado di introdurre un DSB se il PAM è assente17. Da qui, il PAM luoghi un vincolo sulle posizioni genomiche dove una particolare nucleasi di Cas è in grado di fendere. Fortunatamente, nucleasi Cas da diverse specie batteriche in genere presentano diversi requisiti di PAM. Quindi, integrando i vari sistemi di CRISPR-Cas nel nostro toolbox di ingegneria, possiamo espandere la gamma di siti che possono essere mirati in un genoma. Inoltre, un enzima naturale Cas possa essere progettato o si è evoluto per riconoscere sequenze alternative di PAM, ampliando ulteriormente l’ambito di target genomici accessibile a manipolazione18,19,20.
Anche se più sistemi CRISPR-Cas sono disponibili per scopi di ingegneria del genoma, maggior parte degli utenti della tecnologia si affidano principalmente le nucleasi Cas9 da Streptococcus pyogenes (SpCas9) per molteplici ragioni. Innanzitutto, richiede un PAM relativamente semplicemente NGG, a differenza di molte altre proteine di Cas possono fendere solo in presenza di più complesse PAMs. In secondo luogo, è il primo endonucleasi di Cas per essere impiegati con successo in cellule umane21,22,23,24. In terzo luogo, la SpCas9 è di gran lunga l’enzima meglio caratterizzato fino ad oggi. Se un ricercatore vuole utilizzare un altro nucleasi di Cas, è possibile che lui o lei spesso sarebbe poco chiara sul modo migliore per progettare l’esperimento e come ben altri enzimi si esibiranno in diversi contesti biologici rispetto ai SpCas9.
Per fornire chiarezza per le prestazioni relative dei differenti sistemi di CRISPR-Cas, recentemente abbiamo effettuato un confronto sistematico delle cinque endonucleasi Cas – SpCas9, l’enzima Cas9 da Staphylococcus aureus (SaCas9), l’enzima di Cas9 da Neisseria meningitidis (NmCas9), l’enzima di Cas12a da Thermoanaerobacter SP. BV3L6 (AsCas12a) e l’enzima Cas12a dal batterio Lachnospiraceae ND2006 (LbCas12a)25. Per un confronto equo, abbiamo valutato le varie nucleasi Cas utilizzando lo stesso set di siti di destinazione e le altre condizioni sperimentali. I parametri di progettazione di studio anche delineato per ciascun sistema CRISPR-Cas, che dovrebbe servire come un utile riferimento per gli utenti della tecnologia. Qui, per meglio consentire ai ricercatori di fare uso della CRISPR-Cas sistema, forniamo un protocollo dettagliato per l’ingegneria di genoma ottimale con differenti enzimi Cas9 e Cas12a (Vedi Figura 1). Il protocollo comprende non solo i dettagli sperimentali ma anche importanti considerazioni di progettazione per massimizzare la probabilità di un risultato di successo genoma Ingegneria in cellule di mammifero.
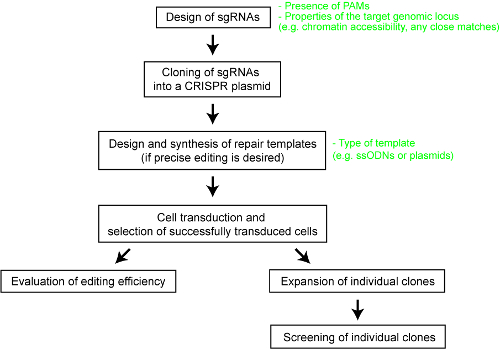
Figura 1 : Una panoramica del flusso di lavoro per generare genoma modificato linee cellulari umane. Clicca qui per visualizzare una versione più grande di questa figura.
