Proteinler ve protein komplekslerinin kristal yapılarının sayısı son yıllarda hızla artmıştır. Bunlar, bu proteinlerin yapısal organizasyon çok değerli anlık mevcut ve yapı-fonksiyon analizi için bir temel sağlar. Ancak, proteinlerin ve bunların fonksiyonları için gerekli olan yapısal değişiklikler, dinamikleri, nadiren X-ışını kristalografisi ile ortaya çıkar. Cryo-elektronmikroskop, diğer taraftan, farklı yapıda protein ve protein kompleksleri yakalamak mümkün ama genellikle ikincil yapı seviye 1 şekilsel değişiklikleri aşağı çözemez. Atomik ayrıntıları, çözelti içinde proteinlerin konformasyonel dinamiği sadece NMR ile çözülebilir, ancak bu yöntem halen göreceli olarak küçük boyutlarda (genellikle ≤ 30 kDa) proteinlere sınırlı ile deneyler engellemektedir proteinlerin yüksek konsantrasyonlarda (≥ 100 uM), ihtiyacı var oligomerizasyon ya da toplama eğilimli proteinler 2. Bir yöntem, kiyüksek çözünürlüklü X-ışını kristalografisi ve kriyo-elektronmikroskop arasında olan ve köprü edebilmektedir protein boyut veya konsantrasyonu ile sınırlı değildir amid hidrojen-1 kütle spektrometrisi (MS) ile bir arada H / 2 H-değişim (HX) 'dir. Son yıllarda bu yöntem, protein dinamiği, protein katlanmasına, protein stabilitesi ve bu şekilsel değişikliklere 3-5 analizi için değerli bir analitik araç geliştirmiştir. Bu yöntemin moleküler temeli protein D 2 O çözeltisi yerleştirildiği zaman, döteryum atomları ile yer değiştirmeye doyurmaya proteinlerde omurga amid hidrojenlerin arasında kararsız doğasıdır. , Zaman içinde protein kütlesindeki artış, daha sonra yüksek çözünürlüklü MS ile ölçülür.
Kısa peptitler yapısal olmayan sadece sıcaklık, katalizör konsantrasyonuna bağlı HX (OH – Şekil 3, 3 H + O, yani pH) ve endüktif, kedi nedeniyle komşu artıkların amino asit yan zincirlerialytic ve sterik etkiler. Iç kimyasal kuru k kanal üzerindeki bu etkiler zarif Bai et al. 6 ile nicelleştirilmiştir edilmiş ve bir program pH ve sıcaklığa bağlı olarak bir polipeptid içinde her bir amino asit için k kanal hesaplar (izniyle Z. Zhang) mevcuttur. Nötr pH ve ortam sıcaklıklarında k ch 10 1 -10 saniye 3 -1 sırasına göre olan. Sıkı bir şekilde katlanmış proteinin iç iyonları kapandı – proteinlerinde HX temel olarak ikincil yapı içinde hidrojen bağı ve OH sulu sınırlı erişimi nedeniyle düşük derecede daha düşük büyüklükte 2-9 emir olabilir. Doğal protein HX bu nedenle kısmen ya da küresel açılımı, kimyasal değişimi ve denkleme göre yerel durumuna yeniden katlama etkisi altına (1) ve k obs açılış hızı k op, kapatma hızı k cl ve içsel kimyasal değişimi bağlıdır gözlenen kur rate k ch denkleme göre (2).
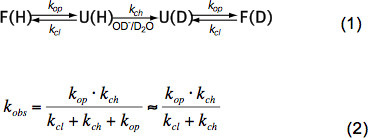
Yerli hal koşullarında k op k ch çok daha küçüktür ve paydada ihmal edilebilir. EX1 ve EX2 denilen iki uç döviz rejimleri vardır. K cl k kanal (EX1) daha küçük ise, gözlenen oranı açma hızına hemen hemen eşit olduğunu ve HX, bir yapı elemanının açılımı hemen gözlem sağlar. , Yapısal elemanın açılması üzerine bir kerede amid proton değişimi, izotop tepe 7 bir bimodal dağılım ile MS kolayca gözlemlenebilir böyle bir değişim rejimi,. K cl k ch çok daha büyük olduğu takdirde orantı sabit katlama açılımı dengesinin eşittir burada, (EX2) gözlenen oranı k ch ile orantılıdır sabiti K u = k op </sub> / K cl. Izotopik dağılımı yaklaşık olarak aynı kalırken, bu koşullar altında, bir çok açılış ve kapanış etkinlik ortalama kütlesi tedrici bir artışa yol açan, döteronların seyahati amid proton değişim önce gereklidir. EX2 rejimi ΔG u açılımı serbest enerji belirlenmesini ve bir yapı elemanının bu nedenle stabilitesini sağlar. Yerli devlet koşul altında EX2 rejimi en yaygın olanıdır. PH ve kaotropik ajanların eklenmesi artış EX1 için değişim mekanizmasını değiştirebilir. Bu nedenle, HX-MS termodinamiği keşfetmek hem de protein katlanma ve yapısal değişikliklerin kinetik parametreler için kullanılabilir.
HX yukarıda belirtildiği gibi doğal olarak pH ve sıcaklığa bağlıdır ve omurga amid grubu bir solvent tamamen açık proton değişim yarılanma ömrü> fizyolojik pH'da 5-400 ms (pH 7.6) ve 30 ° C, fakat 10 dakika arasında olduğu bir pH 2,9> 2 saat ortalama ve 0 ° ile 15 saat(Ca bir yarı ömür. 1-2 dakika ile alışverişi bir polipeptidin, ilk omurga amid bağının proton hariç) C. Bu tür değiş tokuş koşulları altında, yavaş dışarı dahil döteronların içerdiği tüm bilgileri kaybetme ile, bu koşullar altında aktif olan proteazlar (örneğin, pepsin) kullanılarak örnek sindirimi mümkündür. Yavaş alışverişi koşullar altında peptik sindirim tanıtılmasından bu yana, tam uzunlukta proteinlerin genel HX kinetik değil, sadece analiz edilebilir ama HX belirli bölgelerde 8,9 lokalize olabilir. Uzamsal çözünme anda 10-30 kalıntıları arasındaki genel olarak üretilen peptik parçalarının, büyüklüğü ile sınırlıdır. Bununla birlikte, pepsin ile bölünmesi nedeniyle spesifik olmayan doğası oluşturulan üst üste binen parçaları, uzamsal çözünürlük bir artışa neden olabilir. Buna ek olarak, çeşitli başka proteazlar, söndürme koşullar altında, ancak, daha az verimli 10 pepsin daha aktif olduğu bulunmuştur. Dahası increauzamsal çözünürlük se gibi elektron yakalama ayrışma (ECD), elektron transfer ayrılma (ETD) ve kızıl ötesi multiphoton ayrılma (IRMPD) 11-13 gibi döteryumlanma desen korunmuş yöntemler ile, gaz fazında peptitlerin parçalanması ile ulaşılabilir. Bu teknikler nedeniyle çarpışma kaynaklı ayrılma (CID) tarafından görülmektedir intramoleküler proton göçü ("karıştırma"), en yaygın olarak kullanılan parçalama tekniği bir uzamsal çözünürlük kaybını önler. Ancak, bu yöntemler her bir peptit için optimizasyon ve yine bu nedenle oldukça zordur.
HX-MS viral kapsid montaj 14-17 dahil olmak üzere protein-ligand ve protein-protein etkileşimlerini analiz etmek için kullanılmıştır. Protein açılımı ve sıcaklık kaynaklı konformasyonel değişiklikler 7,18,19 incelenmiştir hem de yeniden katlama. Fosforilasyonu ve tek bir amino asit mutasyon-ilişkili şekilsel değişiklikleri 16,20 ve nükleotidlerinide kaynaklı değişimler 21,22 analiz edilmiştir. Bu nedenle, bu yöntem derleme ve moleküler makineleri dinamiklerini analiz etmek için ideal uygun görünüyor. Kimin mekanizması büyük genel ilgi çekici biri aday, Hsp90 şaperon karmaşıktır.
