An overview of the process to generate ELTs – comprising lung slicing, slice clipping and decellularization, and scaffold repopulation – is presented in Figure 3. The ELTs presented here were cultured using primary rat lung microvascular endothelial cells (see Table of Materials), neonatal rat AEC2s, and lipofibroblast-enriched neonatal rat lung fibroblasts36. AEC2s were freshly isolated via magnetic bead-based sorting as previously described37; alternative isolation protocols have been detailed and discussed elsewhere38,39,40. Purity of isolated rat AEC2s can be assessed via flow cytometry for rat-specific AEC2 surface marker RTII-7041, or via staining of a cytocentrifuged cell sample for RTII-70 or pro-surfactant protein C (pSPC). Rat lung fibroblasts were isolated from postnatal day 7-9 rat pups according to an adaptation of a published protocol42 and used at passage 1-2; alternative isolation protocols have been described elsewhere43,44. Purity of isolated fibroblasts can be assessed via staining of cultured or cytocentrifuged cells for mesenchymal marker vimentin, and lipofibroblast enrichment can be assessed via staining for Oil Red O45.
When the lung tissue is uniformly inflated with agarose, and tissue pieces strategically selected and oriented for slicing so as to maximize total and parenchymal tissue area, one rat lung may yield tissue for >100 alveolar ELTs. Strips of PCLS exhibit sufficient mechanical integrity to be clipped into tissue cassettes with few (<5%) instances of tearing (Figure 3B).
The protocol for decellularizing lung slices is closely based on our previously published whole lung decellularization protocol, which by quantitative proteomics was demonstrated to preserve many ECM components at levels not significantly different from those in native lung22. Decellularized slice scaffolds preserve the native architecture of the alveoli, as viewed by hematoxylin and eosin (H&E) staining (Figure 4A,B) and by phase contrast microscopy (Figure 4C). We typically exclude scaffolds containing large airways or vessels (Figure 4D) or tears, although the former can be included if they are of interest to the researcher. Decellularization leads to a 96% reduction in tissue DNA content as measured by an assay for double-stranded DNA (see Table of Materials; 0.50 µg/mg ± 0.073 µg/mg vs 0.018 µg/mg ± 0.0035 µg/mg in native vs decellularized tissue, respectively, mean ± SEM) (Figure 5A), with no DNA visible by hematoxylin staining (Figure 4B). Histological and immunofluorescent staining of decellularized scaffolds reveals maintenance of ECM proteins collagen, elastin, collagen IV, and laminin with architecture and quantity similar to that in native lung slices (Figure 5B-E). Note that the nuclei of native tissues stain blue/black with trichome (for collagen) and EVG (for elastin) stains. Immunofluorescent staining was performed as described previously, using standard methods for staining tissues37. The antibodies used and their respective concentrations are listed in Table 4.
Successful scaffold repopulation leads to highly cellular ELTs after 7 days, with an alveolar-like repopulation pattern visible by light microscopy (Figure 6A-C). In some cases, with very high cellularity, organoid-type structures may be visible (Figure 6A,B). Unsuccessful tissue seeding can be visualized by phase-contrast microscopy during culture (Figure 6C). After culturing tissue scaffolds with AEC2s, fibroblasts, and endothelial cells, ELTs are densely repopulated with alveolar-like structures comprising all three cell lineages (Figure 6D,E). At day 7 or 8, AEC2s maintain cuboidal morphology and express surfactant protein-B (SPB) and lamellar body protein ABCA3, without evidence of significant differentiation to AEC1s (Figure 6E,F). AEC2s are highly proliferative in ELTs, as demonstrated by 5-ethynyl-2'-deoxyuridine (EdU) incorporation following a 2 h pulse at 10 µM (Figure 6G).
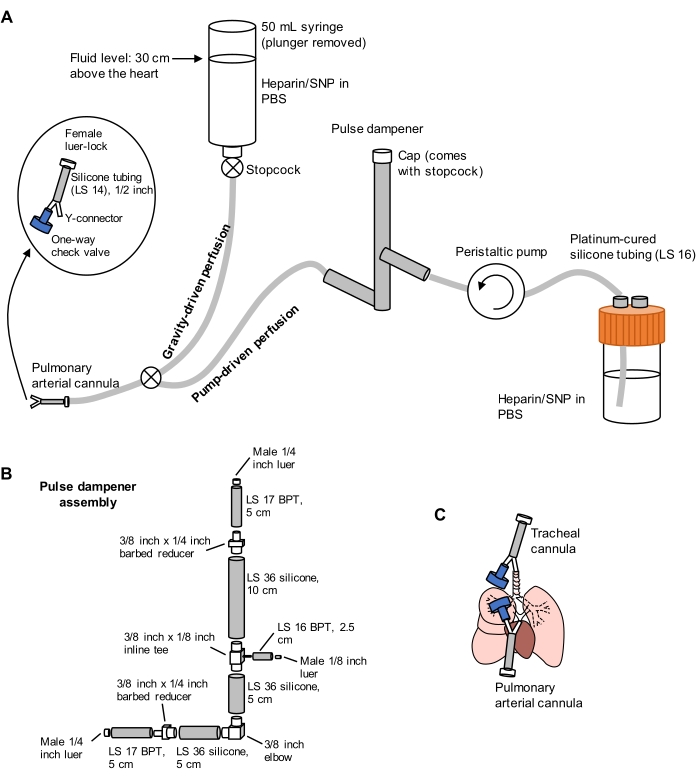
Figure 1: Schematic of perfusion system for lung extraction and clearing. (A) The perfusion system comprises a gravity-driven limb, used for initial cannulation of the pulmonary artery under flow; and a pump-driven limb, used to clear the lungs efficiently after initial cannulation. The pump line includes a "pulse dampener" that dampens the spikes in pressure caused by the pump. The design of the tracheal and pulmonary arterial cannulas is detailed at left. SNP = sodium nitroprusside. (B) Details of pulse dampener assembly. BPT and silicone refer to types of tubing. (C) Positions of tracheal and pulmonary arterial cannulas placed during lung extraction. Please click here to view a larger version of this figure.
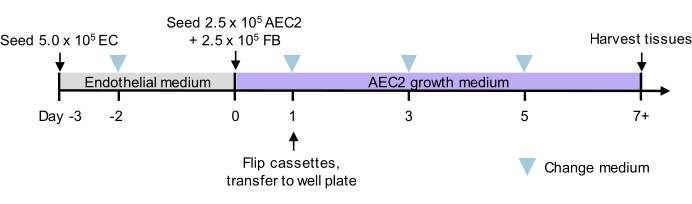
Figure 2: Culture timeline for tri-lineage recellularization. Proposed timeline for tri-lineage ELT seeding and culture, including timing of two-phase seeding. Cell numbers for seeding and culture medium for each phase are indicated. See culture media details in Table 3. AEC2 = alveolar epithelial type 2 cell. EC = endothelial cell. FB = fibroblast. Please click here to view a larger version of this figure.
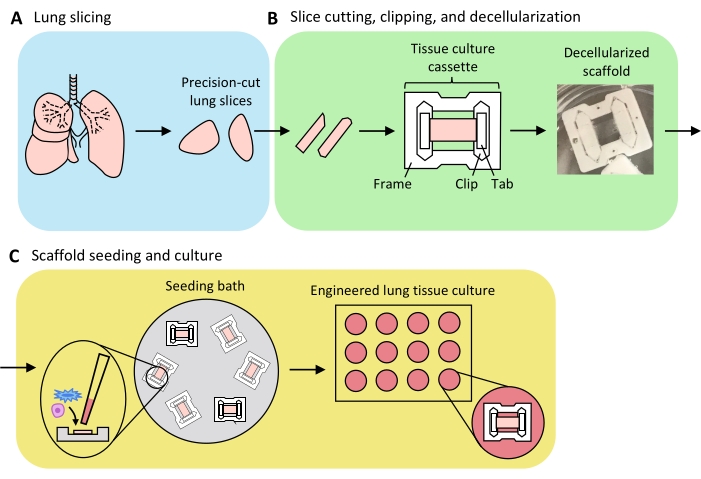
Figure 3: Schematic of engineered lung tissue preparation. (A) Native lung tissue is cut into slices using a vibratome. (B) Precision-cut lung slices are cut into standardized 3 mm wide strips, clipped into polytetrafluorethylene (PTFE) tissue-culture cassettes, and detergent-decellularized to yield acellular extracellular matrix scaffolds. (C) Scaffolds are reseeded in specialized seeding baths that confine the seeding area to the area of the tissue, and then cultured in a standard well plate. Please click here to view a larger version of this figure.
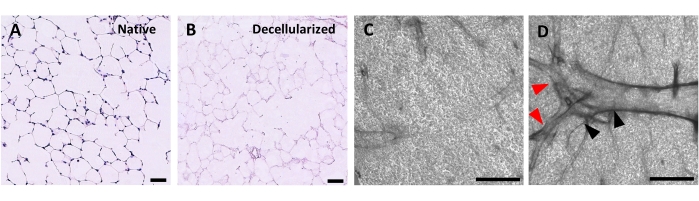
Figure 4: Structure of decellularized lung scaffolds. H&E staining of native (A) and decellularized (B) lung slices showing preservation of alveolar architecture after decellularization. (C,D) Examples of decellularized ECM scaffolds viewed at 5x magnification by phase contrast microscopy, comprising predominately alveolar tissue (C) or containing large branching airways and vessels (D, black and red arrowheads). Scale bars, 50 µm (A,B); 500 µm (C,D). Please click here to view a larger version of this figure.
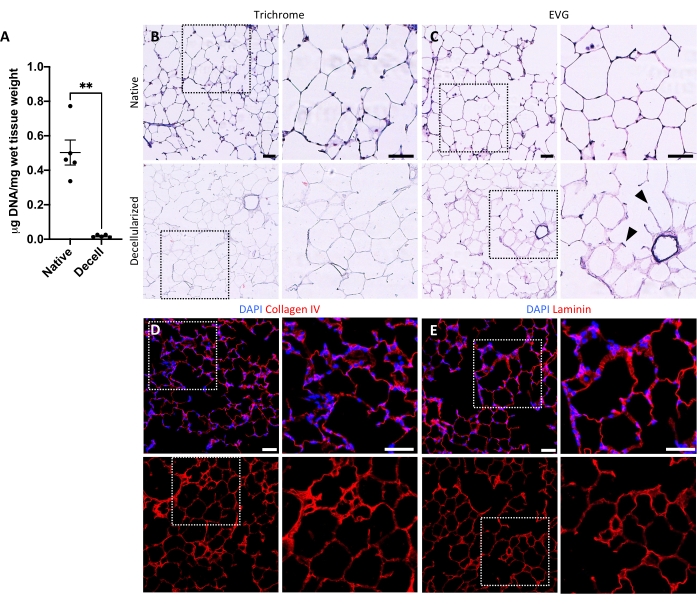
Figure 5: DNA removal and matrix preservation in decellularized lung scaffolds. (A) Quantification of DNA in native and decellularized lung slices (mean ± SEM, n = 5). Welch's t test, **P < 0.01. Decell = decellularized. (B,C) Histological staining of native and decellularized lung slices for collagen (B) and elastin (C). Arrowheads, elastin preserved in alveolar entrance rings of decellularized tissue. (D,E) Immunofluorescent staining of native and decellularized lung slices for collagen IV (D) and laminin (E). Scale bars, 50 µm. In all panels, dotted boxes outline the image region that is magnified to the right in each respective panel. Please click here to view a larger version of this figure.
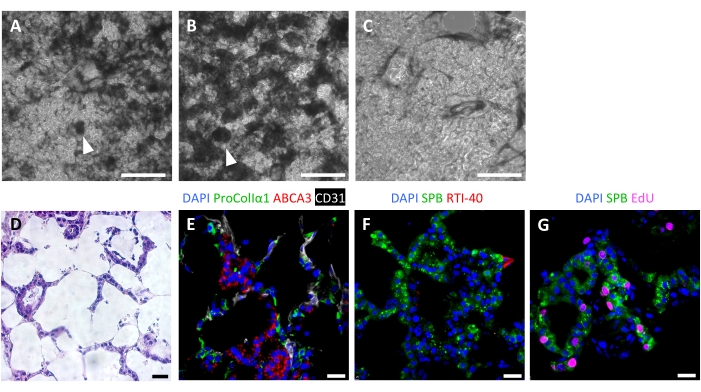
Figure 6: Cellular repopulation of engineered lung tissues. (A-C) Examples of recellularized ELTs on day 7 of culture, as visualized during culture by phase contrast microscopy. The recellularization pattern mirrors the alveolar structure of the tissue. In some areas of high cellularity, organoid-like structures may form (arrowheads). (A) and (B) represent successful cell repopulation, whereas (C) represents a poor level of recellularization after 7 days of culture. (D-G) Staining of recellularized ELTs on day 7 or 8 of culture. (D) H&E staining showing cellular repopulation of the alveolar septa. (E) Immunofluorescent staining labels engrafted proCollagenIα1+ fibroblasts, ABCA3+ AEC2s, and CD31+ endothelial cells. (F) Tissues contain abundant SPB+ AEC2s but few RTI-40 (podoplanin)+ AEC1s under these conditions. (G) Many AEC2s are proliferating in ELTs, as measured by EdU incorporation. Scale bars, 500 µm (A-C); 25 µm (D-G). Please click here to view a larger version of this figure.
Table 1: Decellularization solutions. Preparation details for decellularization solutions. Please click here to download this Table.
Table 2: Decellularization protocol. Details of protocol for decellularizing lung slices. Please click here to download this Table.
Table 3: Culture media. Preparation details for endothelial and AEC2 growth media. Please click here to download this Table.
Table 4: Antibodies used for immunostaining. Details of antibodies and their concentrations used for immunostaining. Please click here to download this Table.
Supplementary File 1: Design for laser cutting tissue culture cassette frames. Please click here to download this File.
Supplementary File 2: Design for laser cutting tissue culture cassette clips. Please click here to download this File.
Supplementary File 3: Design for laser cutting tissue culture cassette tabs. Please click here to download this File.
Supplementary File 4: CAD file for seeding bath mold base. Please click here to download this File.
Supplementary File 5: CAD file for seeding bath mold ring. Please click here to download this File.
