To determine the percentage of mRNA that is ER-associated, COS-7 cells that were transfected with plasmids that contained the placental alkaline phosphatase (ALPP) or cytochrome P450-8B1 (CYP8B1) were either extracted with digitonin and then fixed, or directly fixed (see Figure 1, compare “Unextracted Ctrl” to “Extracted Ctrl”). The non-nuclear fluorescence was quantified in both samples and the fraction of ER-associated mRNA was calculated (Figure 2). Our data clearly shows that for both ALPP and CYP8B1, about 60% of the cytoplasmic mRNA is associated with the ER. Since both of these mRNAs encode proteins that are processed in the ER-lumen, our results are consistent with the idea that such mRNAs are translated on the surface of the ER.
To monitor the ER-association of these transcripts after ribosome dissociation, transfected COS-7 cells were treated with control media, puromycin or HHT for 30 min then extracted, fixed and stained using specific FISH probes directed against each mRNA (for representative images see Figure 1, for quantification of ER- and nuclear-associated mRNA see Figure 3). Note that only ALPP, and not CYP8B1 mRNA, is maintained on the ER after ribosomes are disassembled with puromycin or HHT (Figures 2-3). Importantly the level of nuclear mRNA did not change between the variously treated samples for either mRNA (Figure 3). This constant nuclear FISH signal indicates that the changes in fluorescence intensity in the ER fraction are not due to alterations in mRNA expression or variations in FISH staining.
From these results we conclude that a subset of ER-targeted mRNAs, such as the ALPP transcript, is maintained on the surface of this organelle after ribosome disassembly.
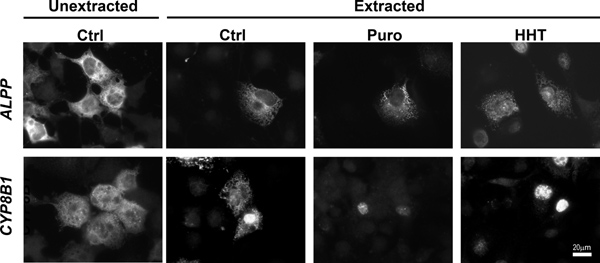
Figure 1. ALPP, but not CYP8B1 mRNA remains associated with the ER independently of ribosomes and translation. COS-7 cells were transfected with plasmids containing either the ALPP (top row), or CYP8B1 (bottom row) genes and allowed to express mRNA for 18-24 hr. The cells were then treated with DMSO control medium (“Ctrl”), puromycin (“Puro”) or HHT for 30 min. Cells were either directly fixed (“unextracted”) or first extracted then fixed. Note that while the control and HHT-treated cells were extracted with digitonin alone, Puro-treated cells were extracted with digitonin and EDTA. Cells were stained for mRNA using specific FISH probes, and imaged. Scale bar = 20 μm.
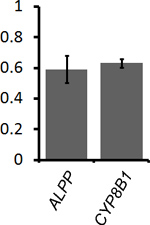
Figure 2. Quantification of the level of ER-association for ALPP and CYP8B1 mRNA. Transfected COS-7 cells were either directly fixed to determine the total level of mRNA in the cytoplasm (see Figure 1, “Unextracted Ctrl” cells) or first extracted and then fixed to determine the amount of ER-associated mRNA (see Figure 1, “Extracted Ctrl” cells). For each experiment the average ER-associated fluorescence of 30-40 extracted cells was divided by the average cytoplasmic fluorescence of 30-40 unextracted cells, to give the fraction of ER-bound mRNA (y-axis). Each bar represents the average value and standard error of three independent experiments.
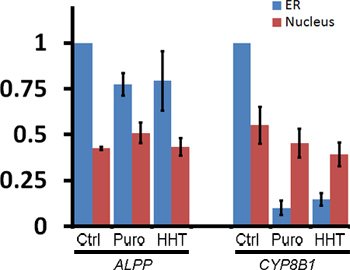
Figure 3. Quantification of the ER-association of ALPP and CYP8B1 mRNA after ribosome dissociation. Transfected COS-7 cells were treated with control medium or various translation inhibitors, extracted, fixed, stained for mRNA using specific FISH probes and imaged (see Figure 1, “Extracted” cells). The fluorescence intensities of mRNA in the ER and nucleus were quantified. Each bar represents the average and standard error of three independent experiments, each consisting of the average integrated intensity of 30 cells over background and normalized to the fluorescence in the ER of control-treated cells (y-axis). Note that although ribosome disruption caused CYP8B1 mRNA to dissociate from the ER, the levels of nuclear mRNA were unaffected.
