Partendo dal modello originale "mosaico fluido" di Singer e Nicolson, l'immagine di membrana plasmatica cellulare è stato costantemente aggiornato nel corso degli ultimi decenni, al fine di comprendere il ruolo emergente del citoscheletro e lipidi domini 1,2.
Le prime osservazioni sono state ottenute da recupero fluorescenza dopo photobleaching (FRAP) svelamento che una frazione significativa di proteine di membrana è immobile 3-5. Questi studi pionieristici, anche se molto informativo, sofferto dalla relativamente scarsa risoluzione spaziale (micron) e di tempo (secondi) di configurazioni FRAP. Inoltre, essendo una misura media insieme, FRAP manca di dare informazioni sul comportamento singola molecola.
In questo contesto, la possibilità di etichettare specificamente una singola molecola di etichetta molto luminosi (permettono lo studio del processo di diffusione una molecola alla volta) ha avuto molto successo. In particolare, spingendo l'risoluzione temporale dell'approccio singola particella di monitoraggio (SPT) per i tempi microsecondi, Kusumi, et al. accesso maturata alle funzioni sconosciute della dinamica dei lipidi e delle proteine che grandemente contribuito al riconoscimento del ruolo di base di actina-membrana scheletro in membrana fisiologia 6 , 7. Questi risultati hanno generato il così chiamato il 'picchetto recinto e' il modello, in cui lipidi e proteine diffusione è regolata da scheletro a base actina-. Tuttavia, al fine di avere accesso alla enorme quantità di informazioni fornite da SPT molte questioni sperimentali devono essere affrontate. In particolare, la procedura di etichettatura è tipicamente composta da molte fasi come la produzione, la purificazione e l'introduzione di specie marcate nel sistema. Inoltre, le grandi etichette, come punti quantici o nanoparticelle metalliche, sono spesso necessari per raggiungere la scala cronologica inferiore al millisecondo e la reticolazione delle molecole bersaglio per l'etichetta non poteva essere evitato in molti casi. Infine, molte traiettoriedevono essere registrati per adattarsi criteri statistici e contemporaneamente è richiesta una bassa densità dell'etichetta per consentire il monitoraggio.
Rispetto a SPT, spettroscopia di fluorescenza di correlazione (FCS), superando molti di questi inconvenienti, rappresenta un approccio molto promettente per studiare la dinamica molecolare. In realtà, FCS funziona bene anche con le etichette dim e densi, consentendo di studiare la dinamica di molecole proteiche-tag fluorescenti nelle cellule trasfettate. Inoltre, permette di raggiungere elevate statistiche in un periodo di tempo limitato. Infine, nonostante la densità "alta" delle etichette FCS fornisce informazioni di singole molecole. Grazie a tutte queste proprietà, FCS rappresenta un approccio molto semplice ed è stato ampiamente applicato a studiare la dinamica dei lipidi e proteine sia in membrane modello e in vivo cellule 8-10. Molti approcci diversi sono stati proposti per aumentare la capacità di FCS per rivelare i dettagli di diffusione molecolare. Per esempio, è stato shproprio che eseguendo FCS sulle aree di osservazione di diverse dimensioni si può definire un "diritto FCS diffusione" caratteristiche illuminanti nascosti del moto molecolare 11,12. Oltre ad essere variato in termini di dimensioni, l'area focale è stato ripetuto anche il 13, si trasferisce in uno spazio lungo le linee 14-20 o coniugati con telecamere veloci 21,22. L'utilizzo di questi correlazione 'spazio-temporale' approcci, parametri biologici rilevanti dei diversi componenti di membrana sono stati quantitativamente descritti su entrambe le membrane modello e quelle biologiche attuali, quindi visione cedimento in membrana organizzazione spaziale.
Tuttavia, in tutti gli FRAP e applicazioni FCS descritto finora la dimensione della zona focale rappresenta un limite di risoluzione spaziale che non può essere superata. Diversi metodi di imaging super-risoluzione sono state recentemente sviluppate per aggirare questo limite. Alcuni sono basati sulla precisione di localizzazione, come ad esempio stocastico la microscopia ottica ricostruzione (STORM) <sup> 23,24, la microscopia localizzazione fotoattivazione (PALM) 25, fluorescenza PALM (FPALM) 26, e single-particelle di tracking PALM (sptPALM) 27: relativamente grande quantità di fotoni necessari ad ogni snapshot, tuttavia, limita la risoluzione temporale di questi metodi per almeno diversi millisecondi, ostacolando così la loro applicabilità in vivo.
Al contrario, una promettente alternativa per Super Resolution Imaging sono stati aperti dal spazialmente modulando l'emissione di fluorescenza con i metodi di deplezione emissione stimolata (STED o reversibili saturabili transizioni ottiche di fluorescenza (RESOLFT)) 28,29. Questi approcci combinano la sagomatura del volume osservazione ben al di sotto del limite di diffrazione con la possibilità di utilizzare microscopi a scansione veloce e sistemi di rilevazione. In combinazione con l'analisi di fluorescenza fluttuazione, microscopia STED permesso di sondare direttamente le dinamiche spazio-temporali nanoscala di lipidi e proteins in membrane cellulari dal vivo 30,31.
Le stesse grandezze fisiche di microscopia basata STED possono essere ottenuti da un spettroscopia immagine correlazione spazio-temporale modificato (STICS 32,33) metodo che è adatto per lo studio della dinamica delle proteine fluorescenti-taggato membrana e / o lipidi in cellule vive e da un microscopio commerciale. Il protocollo sperimentale qui presentata è composta da pochi passi. Il primo richiede l'imaging veloce della regione di interesse sulla membrana. Poi, la pila risultante di immagini viene utilizzato per calcolare le funzioni di correlazione medi spazio-temporali. Con il montaggio della serie di funzioni di correlazione, la 'legge di diffusione' molecolare può essere ottenuto direttamente dalle immagini sotto forma di una diffusività apparente (D app) – vs medi di equipaggio grafico di spostamento. Questa trama dipende in modo critico l'ambiente esplorato dalle molecole e permette di riconoscere direttamente le modalità di diffusione attualidel lipide / proteina di interesse.
In più particolari, come precedentemente indicato 34, la funzione di autocorrelazione spazio-temporale della serie dell'immagine acquisita dipende in modo critico dalla dinamica delle molecole si muovono nella serie di immagini raccolte (si ricorda che lo stesso ragionamento può essere applicato in una linea di acquisizione dove solo una dimensione nello spazio è considerato). In particolare, si definisce la funzione di correlazione come:
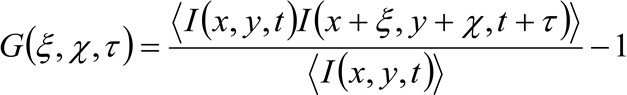 (1)
(1)
dove 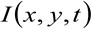 rappresenta l'intensità di fluorescenza misurata nella posizione x, y, al tempo t,
rappresenta l'intensità di fluorescenza misurata nella posizione x, y, al tempo t, ![]() e
e ![]() rappresenta la distanza in x eindicazioni y rispettivamente,
rappresenta la distanza in x eindicazioni y rispettivamente, ![]() rappresenta l'intervallo di tempo, e
rappresenta l'intervallo di tempo, e ![]() rappresenta la media. Questa funzione può essere espressa come:
rappresenta la media. Questa funzione può essere espressa come:
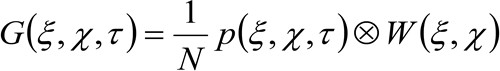 (2)
(2)
dove 'N' rappresenta il numero medio di molecole nella zona di osservazione, ![]() rappresenta l'operazione di convoluzione nello spazio, e
rappresenta l'operazione di convoluzione nello spazio, e 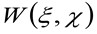 rappresenta l'autocorrelazione della vita strumentale. Quest'ultimo può essere interpretato come una misura di quanto i fotoni di un singolo emettitore sono sparsi nello spazio a causa della configurazione ottica / registrazione (la cosiddetta Point Spread Function, PSF, geneRALLY ben approssimata da una funzione gaussiana). Infine,
rappresenta l'autocorrelazione della vita strumentale. Quest'ultimo può essere interpretato come una misura di quanto i fotoni di un singolo emettitore sono sparsi nello spazio a causa della configurazione ottica / registrazione (la cosiddetta Point Spread Function, PSF, geneRALLY ben approssimata da una funzione gaussiana). Infine, 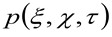 rappresenta la probabilità di trovare una particella a distanza
rappresenta la probabilità di trovare una particella a distanza ![]() e
e ![]() dopo un ritardo
dopo un ritardo ![]() . Se consideriamo una dinamica diffusiva, in cui le particelle si muovono in modo casuale in tutte le direzioni e flussi netti non sono presenti, questa funzione è anche ben approssimata da una funzione gaussiana dove la varianza può essere identificato come il Mean Square Displacement (MSD) della particella in movimento . Così, la vita della funzione di correlazione (indicato anche come
. Se consideriamo una dinamica diffusiva, in cui le particelle si muovono in modo casuale in tutte le direzioni e flussi netti non sono presenti, questa funzione è anche ben approssimata da una funzione gaussiana dove la varianza può essere identificato come il Mean Square Displacement (MSD) della particella in movimento . Così, la vita della funzione di correlazione (indicato anche come ![]() ), Può essere definito come la somma dei DMS particelle e la vita strumentale e può essere misurata con una misura gaussianazione della funzione di correlazione per ogni ritardo. La misurato i MSD può essere usato per calcolare un diffusività apparente delle molecole in movimento
), Può essere definito come la somma dei DMS particelle e la vita strumentale e può essere misurata con una misura gaussianazione della funzione di correlazione per ogni ritardo. La misurato i MSD può essere usato per calcolare un diffusività apparente delle molecole in movimento ![]() e uno spostamento medio
e uno spostamento medio ![]() come:
come:
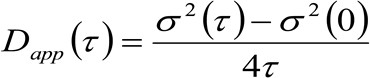 (3)
(3)
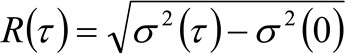 (4)
(4)
Poche considerazioni sul setup sperimentale utilizzato può guidare il lettore attraverso i seguenti sezioni. Al fine di eccitare selettivamente i fluorofori sulla membrana basale delle cellule useremo una riflessione interna (TIR) illuminazione totale vivente, utilizzando una fluorescenza TIR commerciale (TIRF) microscopio (i dettagli si possono trovare nella sezione materiali). Inoltre, al fine di raccogliere the fluorescenza useremo un obiettivo ad alto ingrandimento (100X NA 1,47, alta apertura numerica è necessaria per l'illuminazione TIRF) e una fotocamera EMCCD (dimensione fisica del pixel sul chip 16 micron). Per raggiungere una dimensione di pixel di 100 nm applichiamo una lente di ingrandimento supplementare di 1.6X. Come discusso in seguito, una risoluzione temporale inferiore a 1 msec sarebbe necessario per descrivere correttamente le dinamiche di lipidi di membrana veloci sotto i 100 nm. Per raggiungere questa risoluzione temporale dobbiamo selezionare una regione di interesse (ROI) più piccolo dell'intero circuito integrato della fotocamera (512 x 512). In questo modo, la fotocamera leggerà un numero ridotto di linee aumentando la risoluzione temporale. Tuttavia, in questo regime lettura il lasso di tempo sarebbe limitata dal tempo necessario per spostare le cariche dalla esposizione al chip di lettura sulla fotocamera e di solito dell'ordine di millisecondi per 512 x 512 pixel EMCCD. Per battere questo limite, una tecnologia emergente consente di spostare le ROI linee solo che invece di tutta la struttura, won una riduzione effettiva pratica della dimensione del chip esposto (chiamata modalità ritagliata sensore nel nostro EMCCD). Per questa configurazione sia efficace, il chip di fuori della ROI deve essere coperta da una coppia di feritoie montati nel percorso ottico. Grazie a questa configurazione di risoluzione tempo fino a 10 -4 secondi può essere raggiunto. Si prega di notare, tuttavia, che questo approccio può essere accoppiato con molte diverse configurazioni sperimentali, come spiegato nella sezione 'discussione'.
Dimostrazione del metodo sarà fornito in cellule vive, utilizzando sia un ATTO488 contrassegnata con 1-palmitoil-2-idrossi-sn -glycero-3-fosfoetanolamina (ATTO488-PPE) e una variante di GFP-etichetta della transferrina recettore (GFP TFR). Nel caso di ATTO488-PPE questo approccio può recuperare correttamente un'applicazione D pressoché costante in funzione di spostamento medio indica una diffusione prevalentemente libera, come precedentemente riportato 30,35. Al contrario, TFR-GFP mostra una diminuzione D <sub> app in funzione della cilindrata media, suggerendo diffusione parzialmente confinata 6. Inoltre, in quest'ultimo caso è possibile quantificare la costante diffusione locale e la superficie media di confinamento di numerose micron sul piano membrana.
