A partir del modelo original "mosaico fluido" de Singer y Nicolson, la imagen de la membrana plasmática celular se ha actualizado continuamente durante las últimas décadas con el fin de incluir el papel emergente de citoesqueleto y lípidos dominios 1,2.
Las primeras observaciones fueron obtenidos por la recuperación de fluorescencia después de photobleaching (FRAP) descubrimiento de que una fracción significativa de proteínas de la membrana es inmóvil 3-5. Estos estudios pioneros, aunque muy informativo, sufrieron de la relativamente pobre resolución en el espacio (micras) y el tiempo (segundos) de las configuraciones del FRAP. Además, al ser una medida promedio de conjunto, FRAP le falta en dar información sobre el comportamiento de una sola molécula.
En este contexto, la posibilidad de etiquetar específicamente una única molécula con etiquetas muy brillantes (lo que permite el estudio del proceso de difusión de una molécula a la vez) ha sido muy exitosa. Particularmente, empujando elresolución en el tiempo del enfoque único de rastreo de partículas (SPT) de la escala de tiempo de microsegundos, Kusumi, et al. obtenido acceso a características desconocidas de la dinámica de lípidos y proteínas que contribuyeron en gran medida al reconocimiento del papel de esqueleto de la membrana actina basada en la fisiología de la membrana 6 , 7. Estos resultados generan la llamada el "piquete y la valla" modelo, en el que los lípidos y la proteína está regulada por la difusión esqueleto basado en actina. Sin embargo, con el fin de tener acceso a la enorme cantidad de información proporcionada por SPT muchos temas experimentales que abordar. En particular, el procedimiento de etiquetado se compone típicamente por muchos pasos como la producción, la purificación y la introducción de las especies marcadas en el sistema. Por otra parte, las grandes etiquetas, al igual que los puntos cuánticos o nanopartículas metálicas, a menudo se requieren para llegar a la escala de tiempo sub-milisegundo y la reticulación de las moléculas diana por la etiqueta no se podría evitar en muchos casos. Por último, muchas trayectoriastienen que ser registrado para adaptarse a criterios estadísticos y concomitantemente se requiere una baja densidad de la etiqueta para permitir el seguimiento.
En comparación con SPT, espectroscopia de correlación de fluorescencia (FCS), la superación de muchos de estos inconvenientes, representa un enfoque muy prometedor para estudiar la dinámica molecular. De hecho, FCS también funciona bien con etiquetas tenues y densas, lo que permite estudiar la dinámica de moléculas de proteínas etiquetadas fluorescentes en las células transfectadas transitoriamente. Además, permite alcanzar altas estadísticas en una cantidad limitada de tiempo. Por último, a pesar de la densidad de "alta" de etiquetas FCS proporciona información moléculas individuales. Gracias a todas estas propiedades, FCS representa un enfoque muy sencillo y se ha aplicado ampliamente para estudiar la dinámica de lípidos y proteínas tanto en membranas modelo y en las células vivas 8-10. Se han propuesto muchos enfoques diferentes para aumentar la capacidad de FCS para revelar los detalles de la difusión molecular. Por ejemplo, era shpropia que mediante la realización de FCS en áreas de observación de diferentes tamaños se puede definir una "ley de difusión FCS" características esclarecedoras ocultos de movimiento molecular 11,12. Además de ser variado en tamaño, la esfera de actividad también se duplicó 13, se trasladó en el espacio a lo largo de las líneas 14-20 o conjugados con cámaras rápidas 21,22. El uso de estos correlación 'espacio-temporal' enfoques, parámetros biológicos relevantes de varios componentes de la membrana se designarán cuantitativamente por ambas membranas modelo y las biológicas reales, por tanto, una visión que dé en la membrana organización espacial.
Sin embargo, en toda la FRAP y aplicaciones FCS descrito hasta ahora el tamaño de la esfera de actividad representa un límite de la resolución espacial que no puede ser superada. Varios métodos de imagen super-resolución se han desarrollado recientemente para eludir este límite. Algunos se basan en la precisión de localización, como la microscopía óptica reconstrucción estocástico (TORMENTA) <sup> 23,24, microscopía de localización de fotoactivación (Palm) 25, PALM fluorescencia (FPALM) 26, y de una sola partícula de seguimiento de palma (sptPALM) 27: la cantidad relativamente grande de fotones requeridas en cada instantánea, sin embargo, limita la resolución de tiempo de estos métodos a por lo menos varios milisegundos, lo cual dificulta su aplicabilidad en vivo.
En contraste, una alternativa prometedora para formación de imágenes de súper resolución haber sido abierto por modular espacialmente la emisión de fluorescencia con los métodos de agotamiento de la emisión estimulada (STED o transiciones reversibles saturables ópticos de fluorescencia (RESOLFT)) 28,29. Estos enfoques se combinan la conformación del volumen de observación muy por debajo del límite de difracción con la posibilidad de usar microscopios de barrido rápido y sistemas de detección. En combinación con el análisis de fluctuación de fluorescencia, microscopía STED permitió a la sonda directamente la dinámica espacio-temporales a nanoescala de lípidos y proteins en las membranas celulares en vivo 30,31.
Las mismas cantidades físicas de microscopia basado en STED se pueden obtener por una espectroscopia de correlación de imagen espacio-temporal método modificado (STICS 32,33) que es adecuado para el estudio de la dinámica de las proteínas etiquetadas con fluorescencia de membrana y / o lípidos en células vivas y por un microscopio comercial. El protocolo experimental que se presenta aquí está compuesto por dos pasos. La primera de ellas requiere una imagen rápida de la región de interés en la membrana. Entonces, la pila resultante de imágenes se utiliza para calcular las funciones de correlación promedio espacio-temporales. Mediante el ajuste de la serie de funciones de correlación, la "ley de difusión 'molecular puede ser obtenida directamente de formación de imágenes en forma de una difusividad aparente (D app) – vs -Promedio trazado de desplazamiento. Esta parcela depende de manera crítica sobre el medio ambiente explorado por las moléculas y permite reconocer directamente los modos de difusión realesdel lípido / proteína de interés.
En más detalles, como se indica anteriormente 34, la función de auto-correlación espacio-temporal de la serie de imagen adquirida depende fundamentalmente de la dinámica de las moléculas que se mueven en la serie de imágenes recogido (por favor, tenga en cuenta que el mismo razonamiento se puede aplicar en una adquisición de la línea donde se considera sólo una dimensión en el espacio). En particular, definimos la función de correlación como:
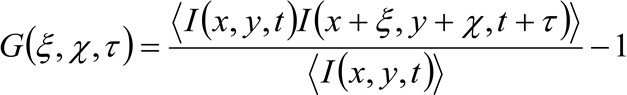 (1)
(1)
donde 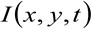 representa la intensidad de fluorescencia medido en la posición x, y y en el tiempo t,
representa la intensidad de fluorescencia medido en la posición x, y y en el tiempo t, ![]() y
y ![]() representa la distancia en la x yY. respectivamente,
representa la distancia en la x yY. respectivamente, ![]() representa el intervalo de tiempo, y
representa el intervalo de tiempo, y ![]() representa la media. Esta función puede expresarse como:
representa la media. Esta función puede expresarse como:
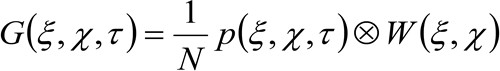 (2)
(2)
donde 'N' representa el número medio de moléculas en el área de observación, ![]() representa la operación de convolución en el espacio, y
representa la operación de convolución en el espacio, y 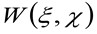 representa la autocorrelación de la cintura instrumental. Este último puede interpretarse como una medida de cómo están distribuidos los fotones de un solo emisor en el espacio debido a la configuración óptica / grabación (el llamado Point Spread Function, PSF, genRally bien aproximada por una función de Gauss). Finalmente,
representa la autocorrelación de la cintura instrumental. Este último puede interpretarse como una medida de cómo están distribuidos los fotones de un solo emisor en el espacio debido a la configuración óptica / grabación (el llamado Point Spread Function, PSF, genRally bien aproximada por una función de Gauss). Finalmente, 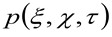 representa la probabilidad de encontrar una partícula a una distancia
representa la probabilidad de encontrar una partícula a una distancia ![]() y
y ![]() después de un tiempo de retardo
después de un tiempo de retardo ![]() . Si tenemos en cuenta una dinámica de difusión, en el que las partículas se mueven al azar en todas las direcciones y los flujos netos no están presentes, esta función también está bien aproximada por una función gaussiana donde la varianza puede ser identificado como el Mean Square Desplazamiento (MSD) de la partícula en movimiento . Por lo tanto, la cintura de la función de correlación (también referido como
. Si tenemos en cuenta una dinámica de difusión, en el que las partículas se mueven al azar en todas las direcciones y los flujos netos no están presentes, esta función también está bien aproximada por una función gaussiana donde la varianza puede ser identificado como el Mean Square Desplazamiento (MSD) de la partícula en movimiento . Por lo tanto, la cintura de la función de correlación (también referido como ![]() ), Se puede definir como la suma de los trastornos musculoesqueléticos de partículas y la cintura instrumental y puede ser medida por un ajuste de GaussTing de la función de correlación para cada retardo de tiempo. La medida i MSD se puede utilizar para calcular una aparente difusividad de las moléculas que se mueven
), Se puede definir como la suma de los trastornos musculoesqueléticos de partículas y la cintura instrumental y puede ser medida por un ajuste de GaussTing de la función de correlación para cada retardo de tiempo. La medida i MSD se puede utilizar para calcular una aparente difusividad de las moléculas que se mueven ![]() y un desplazamiento medio
y un desplazamiento medio ![]() como:
como:
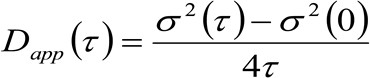 (3)
(3)
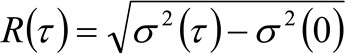 (4)
(4)
Algunas consideraciones sobre la configuración experimental utilizado puede guiar al lector a través de las siguientes secciones. Con el fin de excitar selectivamente los fluoróforos en la membrana basal de las células que utilizaremos una iluminación reflexión interna total (TIR) que viven, utilizando una fluorescencia TIR comercial (TIRF) microscopio (los detalles se pueden encontrar en la sección de materiales). Por otra parte, con el fin de recoger ªe fluorescencia vamos a utilizar un objetivo de gran aumento (100X NA 1,47, se requiere una alta apertura numérica para iluminación TIRF) y una cámara de EMCCD (tamaño físico del píxel en el chip 16 micras). Para llegar a un tamaño de píxel de 100 nm aplicamos una lente de aumento adicional de 1.6X. Como se discute más adelante, una resolución de tiempo por debajo de 1 mseg sería necesario para describir adecuadamente la dinámica de los lípidos de membrana rápido por debajo de 100 nm. Para llegar a esta resolución temporal que necesitamos para seleccionar una región de interés (ROI) más pequeño que todo el chip de la cámara (512 x 512). De esta manera, la cámara leer un número reducido de líneas de aumento de la resolución en el tiempo. Sin embargo, en este régimen de lectura de la hora marco estaría limitado por el tiempo requerido para cambiar los cargos de la exposición a la lectura del chip de la cámara y está normalmente en el orden de milisegundos para EMCCD 512 x 512 píxel. Para superar este límite, una tecnología emergente permite desplazar las líneas de retorno de la inversión sólo en lugar de todo el marco, won una reducción práctica efectiva del tamaño del chip expuesto (llamado Modo Sensor recortada en nuestra EMCCD). Para que esta configuración sea eficaz, el chip fuera de la ROI debe estar cubierto por un par de hendiduras montados en el camino óptico. Gracias a esta configuración una resolución de tiempo hasta 10 -4 segundo se puede lograr. Tenga en cuenta, sin embargo, que este enfoque puede combinarse con diversos montajes experimentales, como se explica en la sección de "discusión".
Demostración del método que se proporcionará en células vivas, usando tanto un marcado ATTO488 1-palmitoil-2-hidroxi-sn-glicero-3 fosfoetanolamina (ATTO488-PPE) y una variante de GFP marcado del receptor de transferrina (GFP TfR). En el caso de ATTO488-PPE este enfoque puede recuperar con éxito una aplicación casi constante D como una función del desplazamiento medio indica una difusión sobre todo libre, como se informó anteriormente 30,35. Por el contrario, TfR-GFP muestra una disminución D <sub> aplicación como una función del desplazamiento promedio, lo que sugiere la difusión parcialmente confinado-6. Además, en el último caso, es posible cuantificar la constante de difusión local y el área promedio de confinamiento durante muchos micras en el plano de la membrana.
