Chicken Recombinant Limbs Assay to Understand Morphogenesis, Patterning, and Early Steps in Cell Differentiation
Summary
Recombinant limbs are a powerful experimental model that allows for studying the process of cell differentiation and the generation of patterns under the influence of embryonic signals. This protocol presents a detailed method for generating recombinant limbs with chicken limb-mesodermal cells, adaptable to other cell types obtained from different organisms.
Abstract
Cell differentiation is the fine-tuned process of cell commitment leading to the formation of different specialized cell types during the establishment of developing tissues and organs. This process is actively maintained in adulthood. Cell differentiation is an ongoing process during the development and homeostasis of organs. Understanding the early steps of cell differentiation is essential to know other complex processes such as morphogenesis. Thus, recombinant chicken limbs are an experimental model that allows the study of cell differentiation and pattern generation under embryonic patterning signals. This experimental model imitates an in vivo environment; it assembles reaggregated cells into an ectodermal cover obtained from an early limb bud. Later, ectoderms are transferred and implanted in a chick embryo receptor to allow its development. This assay was mainly used to evaluate mesodermal limb bud cells; however, it can be applied to other stem or progenitor cells from other organisms.
Introduction
The vertebrate limb is a formidable model for studying cell differentiation, cell proliferation, cell death, pattern formation, and morphogenesis1,2. During development, limbs emerge as bulges from the cells derived from lateral plate mesoderm1. Limb buds consist of a central core of mesodermal cells covered by an ectoderm. From this early structure, a whole and well-formed limb emerge. After the limb bud arises, three axes are recognized: (1) the proximo-distal axis ([PD] shoulder to fingers), (2) the dorso-ventral axis ([DV] from the back of the hand to palm), and (3) the anterior-posterior ([AP] thumb to finger). The proximal-distal axis depends on the apical ectodermal ridge (AER), specialized ectoderm located at the distal tip of the limb bud. The AER is required for outgrowth, survival maintenance, proliferation, and the undifferentiated state of cells receiving signals2,3. On the other hand, the zone of polarizing activity (ZPA) controls anteroposterior patterning4, while the dorsal and ectoderm controls dorsoventral patterning7,8. Integration of three-dimensional patterning implies complex crosstalk between these three axes5. Despite understanding the molecular pathway during limb development, open questions about the mechanisms that control patterning and proper outgrowth to form a whole limb remain unanswered.
Edgar Zwilling developed the recombinant limb (RL) system in 1964 to study the interactions between limb mesenchymal cells and the ectoderm in developing limbs6. The RL system assembles the dissociated-reaggregated limb bud mesoderm into the embryonic limb ectoderm to graft it into the dorsal part of a donor chick embryo. The signals provided by the ectoderm induce the expression of differentiation genes and patterning genes in a spatio-temporal manner, thus inducing the formation of a limb-like structure that can recapitulate the cell programs that occur during limb development7,8,9.
The RL model is valuable for understanding the properties of limb components and the interaction between mesodermal and ectodermal cells6. An RL can be defined as a limb-like structure created by the experimentally assembling or recombining limb bud mesodermal cells inside an ectodermal cover6. The morphogenesis of the RL depends on the characteristics of the mesodermal cells (or other types) that will respond to the ectodermal patterning signals. One of the advantages of this experimental system is its versatility. This characteristic permits the creation of multiple combinations by varying the source of mesodermal cells, such as cells from different developmental stages, from different positions along the limb, or whole (undissociated) or reaggregated cells7,8,9,10. Another example is the capability of obtaining the embryonic ectoderm from species other than chicken, for example, turtle11, quail, or mouse12.
In this sense, the RL technique helps study limb development and the interactions between limb mesenchymal and ectodermal cells from an evolutionary point of view. This technique also has great potential for analyzing the capability of different sources of progenitor cells to differentiate into a limb-like structure by taking advantage of the signals provided by the embryonic ectoderm12,13,14. In contrast to in vitro cultures, the RL permits evaluating the differentiation and morphogenetic potential of a cell population by interpreting embryonic signals from a developing limb9,15.
In this protocol, a step-by-step guide to performing successful RL with reaggregated mesodermal limb bud cells is provided, thus opening the possibility of adapting this protocol with different sources of reaggregated cells or even different ectoderm sources.
Protocol
This research was reviewed and approved by the Institutional Review Board for the Care and Use of Laboratory Animals of the Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México (UNAM, Mexico City, Mexico). A schematic flowchart of the general steps of this protocol is shown in Figure 1A.
1. Embryo incubation and determination of viability
- Incubate fertilized chicken eggs at 38 °C and 60% relative humidity for about three and a half days until they reach the 22 HH stage (according to Hamilton and Hamburger, 1951)16.
NOTE: Freshly fertilized chicken eggs can be stored at 15 °C for up to one week.- To incubate the fertilized eggs, place the eggs vertically with the pointy side down into the humidified incubator. Rotate the eggs during incubation as it is necessary to prevent the developing embryo from adhering to the shell membrane.
NOTE: Fertilized hen eggs can be obtained from local farms. Fertilized White Leghorn chicken eggs are generally used to avoid pigmentation of the feathers in late embryos. Consider incubating enough eggs separately to obtain three structures: (1) limb mesodermal cells, (2) limb ectoderms, and (3) donor embryos to graft the RL.
- To incubate the fertilized eggs, place the eggs vertically with the pointy side down into the humidified incubator. Rotate the eggs during incubation as it is necessary to prevent the developing embryo from adhering to the shell membrane.
- After three and a half days of incubation, remove eggs from the incubator, swab with 70% ethanol, and allow to air dry.
- Identify developing embryos candling the egg to observe the blood vessels and locate the embryo. Discard eggs that do not have an embryo.
NOTE: At this point, distribute the eggs to obtain mesodermal cells, ectoderms, and the hosts for the RL.
2. Obtaining limb mesodermal cells to fill ectoderms
NOTE: Before initiating the manipulations, it is highly recommended to disinfect the working area, the microscopes, and all the instrumental by swabbing with 70% ethanol solution.
- Tap the blunt end of the eggshell with the end of a blunt forceps to open a window and remove about 1 cm x 1 cm of the shell using the forceps.
- Transfer eggs to a plastic or carton holder and placed them one by one under the stereomicroscope. Identify the air membrane and then remove it by picking a small hole where no vessels are found. Pull this area with the aid of fine surgical forceps.
NOTE: The air membrane can be identified as the white and opaque membrane observed immediately after windowing the egg.- Remove any small piece of eggshell that may come into contact with the embryo.
NOTE: Verify that the embryos are in the 22 HH stage before initiating the protocol. Embryos in earlier stages can be returned to the incubator after sealing the eggshell window with tape.
- Remove any small piece of eggshell that may come into contact with the embryo.
- Open the amniotic sac completely by tearing open the amnion using the fine surgical forceps (see Table of Materials).
NOTE: Amnion can be identified as a transparent membrane that closely covers the embryo and is filled with amniotic fluid. - Carefully remove the embryo from the egg using a pair of blunt forceps and then transfer into a sterile Petri dish containing ice-cold phosphate-buffered saline solution (1x PBS). Repeat this step for the remaining eggs. Consider that only ~8-10 embryos are sufficient to obtain the mesodermal cells.
- Wash the embryos once in ice-cold 1x PBS, and with the aid of a stereomicroscope (see Table of Materials), withdraw any remaining membranes. Locate the hindlimb buds.
NOTE: Limb buds are rounded structures situated along the anterior-posterior axis of the embryo as protrusions from the flank. The hindlimbs are located more posteriorly than the forelimbs. - Maintain the embryos in clean 1x PBS, and using a pair of fine surgical forceps, snip each hindlimb bud longitudinally. Cut the bud very close to the flank of the embryo to dissect out the whole hindlimb buds. Repeat this step with the remaining embryos.
- Use a plastic transfer pipette to transfer the limb buds into an empty 1.5 mL microcentrifuge tube.
- Use a pipette tip to remove any excess of 1x PBS and replace PBS with 500 µL of 0.5% trypsin solution (see Table of Materials). Incubate limb buds in a thermoblock for 7 min at 37 °C.
- Remove the trypsin solution and replace it with 500 µL of collagenase type IV at 2 mg/mL in Hanks Balanced Salt Solution (see Table of Materials), then incubate it in a thermoblock at 37 °C for 8 min.
NOTE: Trypsin incubation slightly detaches the ectoderms, while collagenase disaggregate mesodermal cells. - After incubation, remove as much collagenase as possible and replace it with 1 mL of cold Dulbecco's Modified Eagle Medium-high glucose (DMEM-HG) medium supplemented with 10% fetal bovine serum (FBS) to inactivate the enzymes. Pipette the mixture gently ~10 times.
- Filter the suspension containing the cells with a cell strainer of 70 µm to leave behind the ectoderms.
NOTE: Filtration will also remove any undigested limb buds, leaving behind a suspension of single cells. - After filtering, pipette the suspension again 5-10 times and centrifuge at 200 x g for 5 min at room temperature, then discard the supernatant.
- Use 1 ml of DMEM-HG supplemented with 10% FBS to wash the excess collagenase. Centrifuge the suspension at 200 x g for 5 min at room temperature.
- Discard the medium carefully by pipetting and then replace it with 1 mL of fresh DMEM-HG supplemented with 10% FBS without disturbing the cells in the bottom of the tube.
- Allow the cells to form a compact pellet (reaggregate) after incubating them for 1-1.5 h at 37 °C in a thermoblock.
NOTE: While mesodermal cells are forming the pellet, it is convenient to obtain the limb ectoderms.
3. Obtaining the limb ectoderms
- Repeat steps 1-6 from section 2 separately with the other 22 HH chicken embryos chosen to obtain the ectoderms.
NOTE: The number of embryos for this purpose is proportional to the final number of the RL desired. A ratio of 2:1 ectoderms-RL is appropriate. - After obtaining the hindlimb buds, use a plastic pipette to transfer them into an empty microcentrifuge tube. Remove any excess 1x PBS and replace it with 500 µL of 0.5% trypsin in sterile 1x PBS. Incubate the mixture for 30 min at 37 °C in a thermoblock.
- After the enzymatic digestion, transfer the trypsin solution containing the limb buds into a sterile Petri dish.
- Remove the excess trypsin as much as possible using a micropipette; flood the Petri dish with ice-cold 1x PBS supplemented with 10% of FBS until all limb buds are covered.
NOTE: FBS can be substituted for horse serum. - Identify the limb buds under the stereomicroscope; identify the ectoderm as a slightly detached transparent membrane from the limb mesodermal cells (Figure 1B).
- Hold the most proximal end of the limb bud with the aid of fine surgical forceps while carefully detach and separate the ectoderm layer using the other forceps.
NOTE: Obtaining the ectoderms could be difficult due to mesodermal cell clumping and attaching to ectoderms, thus causing them to become sticky. It is recommended to constantly discard the mesodermal cells to maintain only the ectoderms in the Petri dish. - Maintain the ectoderms in ice-cold 1x PBS-10% FBS solution.
NOTE: Ectoderms can be stored at 4 °C in case the reaggregated mesoderm is not ready. However, it is preferable to coordinate the incubation time of the pellet with the ectodermal obtaining.
4. Assembling mesodermal cells inside the ectodermal cover
NOTE: For this, it is necessary to have the empty ectoderms in a Petri dish with sterile 1x PBS-10% FBS solution containing a formed pellet of mesodermal cells.
- After the pellet is formed (see section 2), discard ~600 µL of the medium from the tube by pipetting.
- Carefully detach the pellet from the bottom using a pipette tip and without smashing or destroying it.
- When the pellet is completely detached from the bottom of the tube, turn the tube upside down to transfer the pellet into the Petri dish containing the empty ectoderms (Figure 1C).
- Cut a small piece of the pellet with the aid of a pair of fine surgical forceps and place it as close as possible to the ectoderm.
- As if it were a bag, open the ectoderm with the surgical forceps, and place the piece of pellet as tightly as possible into the ectodermal cover. Repeat this step for as many RLs as desired (Figure 1D).
NOTE: Cut the pieces of the pellet one by one, and fill the ectoderms to maintain clean the 1x PBS-10% FBS solution. The size of the pellet needs to correspond to each ectoderm. - Allow the ectoderm and the mesoderm to heal together for ~30 min at room temperature and then graft them into the embryo host. Discard the unused ectoderm and mesoderm.
5. Transplantation of the filled ectoderm into a host embryo
NOTE: Before transplanting the ectoderms, arrange two stereomicroscopes next to each other on a benchtop, one for embryo manipulation and RL grafting. The other is for maintaining the filled ectoderms ready to transfer into the embryo.
- Select the number of desired eggs to graft the filled ectoderms.
- Using the end of a blunt forceps, tap the eggshell of the 22 HH host embryos to open a window. Identify the air membrane and, using a pair of fine surgical forceps, remove it completely.
- Open the amniotic sac near the forelimb to expose the right flank of the embryo. Open only the amount needed to accomplish the procedure.
- Guided by the forelimb position, perform wound scratching with a tungsten needle (see Table of Materials) to the length of 2-3 somites, slightly damaging the mesoderm until it bleeds.
- Individually transfer a filled ectoderm (RL) into the chick embryo and place the base of the recombinant limb over the somite wound.
NOTE: RL transfer can be done with a plastic pipette or with an angled slit knife. - Fix the RL correctly with two pieces of palladium wire of 0.025 mm diameter and 0.5-1 mm length (see Table of Materials). Align the base of the RL with the wound to ensure that it will be attached to the flank of the host embryo and vascularized (Figure 1E).
- Seal the window with tape, and return the egg to the incubator. Collect the embryos at the desired time points for analysis.
Representative Results
Recognizing a well-performed recombinant limb
After grafting, the manipulated embryos were returned to the incubator to allow the RL to develop. The incubation time correlated with the requirements of the experiment. Nevertheless, the RL can be easily distinguished after 12 h of implantation. To determine whether the implantation was adequate, the RL was observed as a protuberance that was securely attached to the mesodermal wall of the donor embryo (Figure 2A). On the contrary, whether either cell viability and/or the graft failed, the RL was detached from the mesodermal wall or presented a rough morphology (Figure 2B).
Morphological and patterning examination of recombinant limbs
For morphological examination, RL was stained with Alcian blue17 to observe the formation of skeletal elements and their patterning. It is recommended to stain the whole trunk of the donor embryo to avoid missing the RL during the procedure (Figure 3A). Alternatively, before clearing the RL, images in ethanol solution were obtained to observe the morphology of the RL or perform quantitative measurements (Figure 3B). Stained or unstained RL was sliced to observe tissue structure or identify cell type (Figure 3C).
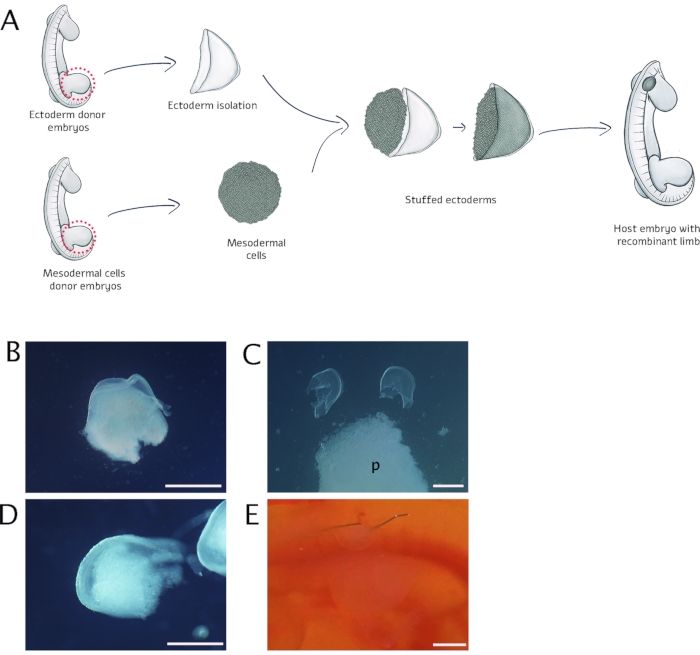
Figure 1: Schematic representation of the experimental design of recombinant limbs (RL). (A) RL was performed by assembling limb bud mesoderm from a donor 22 HH chick embryo inside an ectodermal cover obtained from another 22 HH chick embryo donor. Ectoderms were tightly stuffed with mesodermal cells. After assembly, stuffed ectoderms were transferred and fixed with palladium wires on top of a previous somite's wound. (B) A limb bud with its ectoderm detached after trypsin treatment. (C) The pellet was obtained after its formation near to the empty ectoderms, ready to be filled. (D) An ectodermal cover filled with mesodermal cells is shown. (E) Fixing the RL in the host embryo with palladium wires. Please note that the RL was positioned on the embryo's right flank near the forelimb bud; p: pellet. Scale bar = 500 µm. Please click here to view a larger version of this figure.
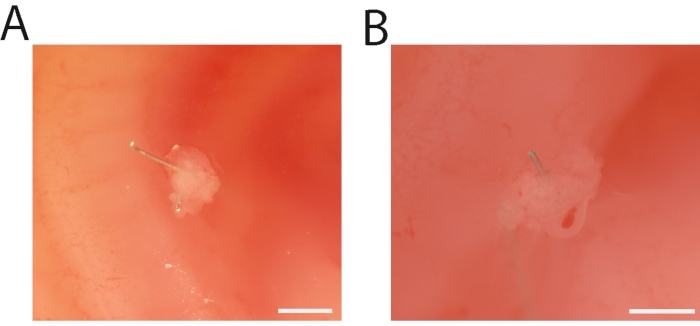
Figure 2: Freshly obtained chicken-chicken recombinant limbs. (A) A 24 h RL attached to the mesoderm wall is shown. (B) An unsuccessful 24 h RL is shown. Please note that the palladium wire is not fixing the RL; consequently, the RL detached from the mesodermal wall and presented a rough morphology. Scale bar = 500 µm. Please click here to view a larger version of this figure.
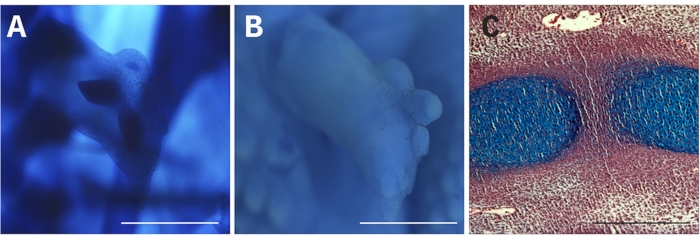
Figure 3: Morphological analysis of recombinant limbs. (A) Alcian blue staining to demonstrate skeletal elements in a 6-day chicken-chicken RL. (B) The same RL was shown in (A) before clearing the Alcian blue. (C) Sagittal slice of an RL stained with Alcian blue staining and with hematoxylin and eosin. Scale bar = 100 µm. Please click here to view a larger version of this figure.
Discussion
In general, the RL protocol can be divided into five steps: (1) embryo incubation, (2) obtaining limb mesodermal cells to fill the ectoderms, (3) obtaining the ectoderms, (4) assembling mesodermal cells inside the ectodermal covers, and (5) transplantation of the filled ectoderms into the host embryos. The major limitation of the RL technique is the long, detailed protocol, which has many critical points that require patience to perform appropriately. To successfully complete the protocol, critical moments need to be identified. During mesodermal cell procurement, the integrity and viability of the cells are essential. Cell death will prevent proper RL development. In a similar vein, correct ectoderm manipulation is necessary to guarantee the interaction between mesodermal and ectodermal cells. When ectoderms are stuffed, mesodermal cells must be as close as possible to the distal ectoderm beneath the AER. For both mesodermal and ectodermal procurement, the developmental stage of the donor embryos is also critical. It must be considered that the mesodermal cells will respond differentially according to their developmental stage. However, the developing stage can be freely selected according to the experimental requirements. Still, the 22 HH stage needs to be maintained to make obtaining ectoderms easier, thus maintaining cellular integrity and signaling. Finally, good grafting and fixing are essential for ensuring correct RL integration to the embryo wall and its development.
Edgar Zwilling first reported the RL system in 19646, after which many research groups implemented it to answer several interesting biological questions. The protocol of the RL has been previously described in length by Marian Ros et al. as a standard method to manipulate the developing chick limb bud18, which explains other ways to window the eggs and perform RL with whole wing or leg and to perform RL with reaggregated mesoderm. However, some variations between the Ros. et al. protocol and the present protocol described here can be found. In their protocol, limb buds from embryos as ectoderm donors are incubated with trypsin in ice-cold PBS for ~2 h. After initiating ectoderm incubation, they immediately obtained limb buds from embryos as mesoderm donors, then fragmented the limb buds, digested them, and removed the ectoderm manually to form the mesodermal pellet after incubating for 30 min. Here, first, the limb buds were obtained from embryos to be used as mesodermal donors, after which the whole limb bud is digested by incubating with trypsin and collagenase. Ectoderms are then removed by filtration, and the pellet is incubated between 1-1.5 h. The advantage of this method to obtain the mesodermal pellet is that the treatment with trypsin detaches the intact ectoderms from the mesoderm while the collagenase treatment digests mesodermal cells. Therefore, it is possible to filter the ectodermal tissue and discard it. On the other hand, more pellet incubation time allows it to compact better, which helps when ectoderms fill. Another difference between the two protocols is that Ros et al. peeled off the ectoderms one by one and transferred them to the Petri dish containing the pellet. In contrast, all the ectoderms are dissected and collected in a Petri dish in the present protocol. The pellet is transferred to the Petri dish to fill the ectoderms. By following this method, the ectoderms can be prepared simultaneously with grafting. As with many other protocols, how RLs are performed may vary; however, the steps in both protocols adequately describe the critical stages of the technique to produce a successful manipulation.
Previous work has demonstrated that dissociated polarizing zone cells inhibit morphogenesis when randomly dispersed among mesoderm in RL7,10. Thus, it is optional to eliminate cells from the ZPA before forming the mesodermal pellet. Later, the ZPA cells (or sonic hedgehog embedded beads) can be used to induce the RL to develop A-P polarity8,9,19.
The RL experimental model is adaptable to a variety of scenarios. Recombination can be implemented with limb cells from different developmental or mature (fore- or hindlimb) stages, other positions along the three limb axes, and with dissociated-reaggregated cells or undissociated-fragmented mesoderm15. Interestingly, previous studies have reported using the RL model to study the behavior of different combinations of mutant and wild-type mesoderm and ectoderms13,14,20,21 or using electroporated cells22.
Considering that limb development is an evolutionarily conserved process, the ectoderm sources also can vary from the chicken, quail, duck, mouse, or rat ectoderms can be obtained following the same described protocol. Another possibility is changing the mesodermal -or even other cell types or sources to produce interspecies RL.
In conclusion, RL is a phenomenal model to study morphogenesis, patterning, cell-cell interactions, cell migration, and cell differentiation at the cellular and molecular levels. Because the procedure of RL allows multiple variations, it permits potential applications across numerous biological questions without being restricted to chicken-limb developmental biology.
Divulgations
The authors have nothing to disclose.
Acknowledgements
We thank to Estefania Garay-Pacheco for images in Figure 2 and to Maria Valeria Chimal-Montes de Oca for artwork. This work was supported by the Dirección General de Asuntos del Personal Académico (DGAPA)-Universidad Nacional Autónoma de México [grant numbers IN211117 and IN213314] and Consejo Nacional de Ciencia y Tecnología (CONACyT) [grant number 1887 CONACyT-Fronteras de la Ciencia] awarded to JC-M. JC M-L was the recipient of a postdoctoral fellowship from the Consejo Nacional de Ciencia y Tecnología (CONACyT-Fronteras de la Ciencia-1887).
Materials
| Alcian Blue 8GX | Sigma | A5268 | |
| Angled slit knife | Alcon | 2.75mm DB | |
| Blunt forceps | Fine Science Tools | 11052-10 | |
| Collagenase type IV | Gibco | 1704-019 | |
| DMEM-HG | Sigma | D5796 | |
| Egg incubator | Incumatic de Mexico | Incumatic 1000 | |
| Fetal Bovine Serum | Gibco | 16000069 | |
| Fine surgical forceps | Fine Science Tools | 9115-10 | |
| Hanks Balanced Salt Solution | Sigma | H6648 | |
| Microcentrifuge | Eppendorf | 5417R | |
| Micropipet | NA | NA | |
| Palladium wire | GoodFellow | 7440 05-3 | |
| Petri dish | Nest | 705001 | |
| Pippette | crmglobe | PF1016 | |
| Stereomicroscope | Zeiss | Stemi DV4 | |
| Tape | NA | NA | |
| Trypsin porcine | Merck | 9002 07-7 | |
| Tungsten needle | GoodFellow | E74-15096/01 |
References
- Malashichev, Y., Christ, B., Pröls, F. Avian pelvis originates from lateral plate mesoderm and its development requires signals from both ectoderm and paraxial mesoderm. Cell and Tissue Research. 331 (3), 595-604 (2008).
- Mahmood, R., et al. A role for FGF-8 in the initiation and maintenance of vertebrate limb bud outgrowth. Current Biology. 5 (7), 797-806 (1995).
- Yu, K., Ornitz, D. M. FGF signaling regulates mesenchymal differentiation and skeletal patterning along the limb bud proximodistal axis. Development. 135 (3), 483-491 (2008).
- Riddle, R. D., Johnson, R. L., Laufer, E., Tabin, C. Sonic hedgehog mediates the polarizing activity of the ZPA. Cell. 75 (5), 1401-1416 (1993).
- McQueen, C., Towers, M. Establishing the pattern of the vertebrate limb. Development. 147 (17), (2020).
- Zwilling, E. Development of fragmented and of dissociated limb bud mesoderm. Developmental biology. 9 (1), 20-37 (1964).
- Frederick, J. M., Fallon, J. F. The proportion and distribution of polarizing zone cells causing morphogenetic inhibition when coaggregated with anterior half wing mesoderm in recombinant limbs. Development. 67 (1), 13-25 (1982).
- Ros, M. A., Lyons, G. E., Mackem, S., Fallon, J. F. Recombinant limbs as a model to study homeobox gene regulation during limb development. Biologie du développement. 166 (1), 59-72 (1994).
- Piedra, M. E., Rivero, F. B., Fernandez-Teran, M., Ros, M. A. Pattern formation and regulation of gene expressions in chick recombinant limbs. Mechanisms of Development. 90 (2), 167-179 (2000).
- Crosby, G. M., Fallon, J. F. Inhibitory effect on limb morphogenesis by cells of the polarizing zone coaggregated with pre-or postaxial wing bud mesoderm. Biologie du développement. 46 (1), 28-39 (1975).
- Fallon, J. F., Simandl, B. K. Interactions between chick limb bud mesoderm and reptile ectoderm result in limb outgrowth in the limbless mutant. Anatomical Record. 208, 53-54 (1984).
- Kuhlman, J., Niswander, L. Limb deformity proteins: role in mesodermal induction of the apical ectodermal ridge. Development. 124 (1), 133-139 (1997).
- Goetinck, P. F., Abbott, U. K. Studies on limb morphogenesis. I. Experiments with the polydactylous mutant, talpid. Journal of Experimental Zoology. 155, 161-170 (1964).
- Carrington, J. L., Fallon, J. F. Initial limb budding is independent of apical ectodermal ridge activity; evidence from a limbless mutant. Development. 104 (3), 361-367 (1988).
- Fernandez-Teran, M., Piedra, M. E., Ros, M. A., Fallon, J. F. The recombinant limb as a model for the study of limb patterning, and its application to muscle development. Cell and Tissue Research. 296 (1), 121-129 (1999).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88 (1), 49-92 (1951).
- Ganan, Y., Macias, D., Duterque-Coquillaud, M., Ros, M. A., Hurle, J. M. Role of TGF beta s and BMPs as signals controlling the position of the digits and the areas of interdigital cell death in the developing chick limb autopod. Development. 122 (8), 2349-2357 (1996).
- Ros, M. A., Simandl, B. K., Clark, A. W., Fallon, J. F. Methods for manipulating the chick limb bud to study gene expression, tissue interactions, and patterning. Developmental Biology Protocols. 137, 245-266 (2000).
- MacCabe, J. A., Saunders, J. W., Pickett, M. The control of the anteroposterior and dorsoventral axes in embryonic chick limbs constructed of dissociated and reaggregated limb-bud mesoderm. Biologie du développement. 31 (2), 323-335 (1973).
- Zwilling, E. Effects of contact between mutant (wingless) limb buds and those of genetically normal chick embryos: confirmation of a hypothesis. Biologie du développement. 39 (1), 37-48 (1974).
- Prahlad, K. V., Skala, G., Jones, D. G., Briles, W. E. Limbless: A new genetic mutant in the chick. Journal of Experimental Zoology. 209 (3), 427-434 (1979).
- Marin Llera, J. C., Lorda-Diez, C. I., Hurle, J., Chimal-Monroy, J. SCA-1/Ly6A mesodermal skeletal progenitor subpopulations reveal differential commitment of early limb bud cells. Frontiers in Cell and Developmental Biology. 9, 656999 (2021).
