يقدم علم البصريات الوراثي، وهو التحكم في العمليات البيولوجية باستخدام البروتينات المستجيبة للضوء، استراتيجية جديدة للتحكم ديناميكيا في التخمير الميكروبي لإنتاج المواد الكيميائية والبروتين1،2. غالبا ما يضعف عبء المسارات الأيضية الهندسية وسمية بعض المواد الوسيطة والمنتجات نمو الخلايا3. ويمكن أن تؤدي هذه الضغوط إلى ضعف تراكم الكتلة الأحيائية وانخفاض الإنتاجية3. ويمكن التصدي لهذا التحدي عن طريق تقسيم عمليات التخمير مؤقتا إلى مرحلة نمو وإنتاج، تكرس موارد التمثيل الغذائي لتراكم الكتلة الحيوية أو توليف المنتجات على التوالي4. لقد أظهرنا مؤخرا أن الانتقال من النمو إلى الإنتاج في هذا التخمير ثنائي المراحل يمكن أن يحدث مع التغيرات في ظروف الإضاءة5،6،7. توفر القابلية العالية للضبط والانعكاس والتعامد لمدخلات الضوء8 مزايا فريدة للتخمير الذي يتم التحكم فيه بالضوء والذي يصعب أو يستحيل تكراره مع المحفزات الكيميائية المستخدمة في التحكم الديناميكي في التخمير التقليدي ثنائي الطور4،9،10،11.
تم استخدام بروتين EL222 المستجيب للضوء الأزرق المشتق من Erythrobacter litoralis لتطوير العديد من الدوائر البصرية الجينية للهندسة الأيضية في Saccharomyces cerevisiae5,7,12,13. يحتوي EL222 على مجال مستشعر جهد الضوء والأكسجين (LOV) الذي يخضع لتحول توافقي عند تنشيط الضوء الأزرق (465 نانومتر) ، مما يسمح له بالارتباط بتسلسل الحمض النووي المتشابه (C120)13. يؤدي دمج EL222 مع مجال تنشيط VP16 الفيروسي (VP16-EL222) إلى عامل نسخ مستجيب للضوء الأزرق يمكنه تنشيط التعبير الجيني بشكل عكسي في S. cerevisiae7 والكائنات الحية الأخرى14 من المروج الاصطناعي PC120. تم تطوير العديد من الدوائر القائمة على EL222 واستخدامها للإنتاج الكيميائي في S. cerevisiae ، مثل نظام OptoEXP الأساسي المنشط بالضوء 7 ، حيث يتم التعبير عن الجين محل الاهتمام مباشرة من PC120 (الشكل 1A). ومع ذلك ، فإن المخاوف من اختراق الضوء في كثافات الخلايا العالية التي تصادف عادة في مرحلة إنتاج التخمير حفزتنا على تطوير دوائر مقلوبة يتم تحفيزها في الظلام ، مثل دوائر OptoINVRT و OptoQ-INVRT (الشكل 1B) 5،7،13. تسخر هذه الأنظمة الجالاكتوز (GAL) أو حمض الكينيك (Q) من S. cerevisiae و N. crassa ، على التوالي ، وتتحكم في مثبطاتها المقابلة (GAL80 و QS) باستخدام VP16-EL222 ، لقمع التعبير الجيني في الضوء وحثه بقوة في الظلام. يؤدي الجمع بين دوائر OptoEXP و OptoINVRT إلى التحكم ثنائي الاتجاه في التعبير الجيني ، مما يتيح التخمير ثنائي الطور الذي يتم فيه تحفيز مرحلة النمو بالضوء الأزرق ، ومرحلة الإنتاج مع الظلام (الشكل 2A)5,7.
إن استخدام الضوء بدلا من الظلام للحث على التعبير الجيني خلال مرحلة الإنتاج من شأنه أن يوسع بشكل كبير قدرات الضوابط البصرية الجينية ولكنه سيتطلب أيضا التغلب على قيود اختراق الضوء لكثافات الخلايا العالية التي تصادف عادة في مرحلة التخمير هذه. وتحقيقا لهذه الغاية، قمنا بتطوير دوائر، تعرف باسم OptoAMP و OptoQ-AMP، تعمل على تضخيم الاستجابة النسخية لتحفيز الضوء الأزرق. تستخدم هذه الدوائر طفرات من النوع البري أو شديد الحساسية من VP16-EL222 للتحكم في إنتاج منشطات النسخ Gal4p أو QF2 من تنظيمات GAL أو Q ، على التوالي ، مما يحقق حساسية معززة وتعبيرا جينيا أقوى مع الضوء12,13 (الشكل 1C). يمكن لدوائر OptoAMP تحقيق تحريض ضوئي كامل ومتجانس في مفاعلات حيوية سعة 5 لتر بكثافة بصرية (تقاس عند 600 نانومتر; OD600) قيم لا تقل عن 40 مع ~ 0.35٪ فقط من الإضاءة (5٪ جرعة ضوئية على ~ 7٪ فقط من السطح السائب). وهذا يدل على درجة أعلى من الحساسية مقارنة ب OptoEXP، الذي يتطلب إضاءة بنسبة 100٪ تقريبا12. إن القدرة على تحفيز التعبير الجيني بفعالية مع الضوء عند كثافة الخلايا العالية تفتح فرصا جديدة للتحكم الديناميكي في عمليات التخمير. ويشمل ذلك عمليات التخمير التشغيلية في أكثر من مرحلتين زمنيتين، مثل التخمير ثلاثي المراحل، حيث يتم إنشاء مراحل النمو والحث والإنتاج بجداول إضاءة فريدة لتحسين الإنتاج الكيميائي (الشكل 2B)12.
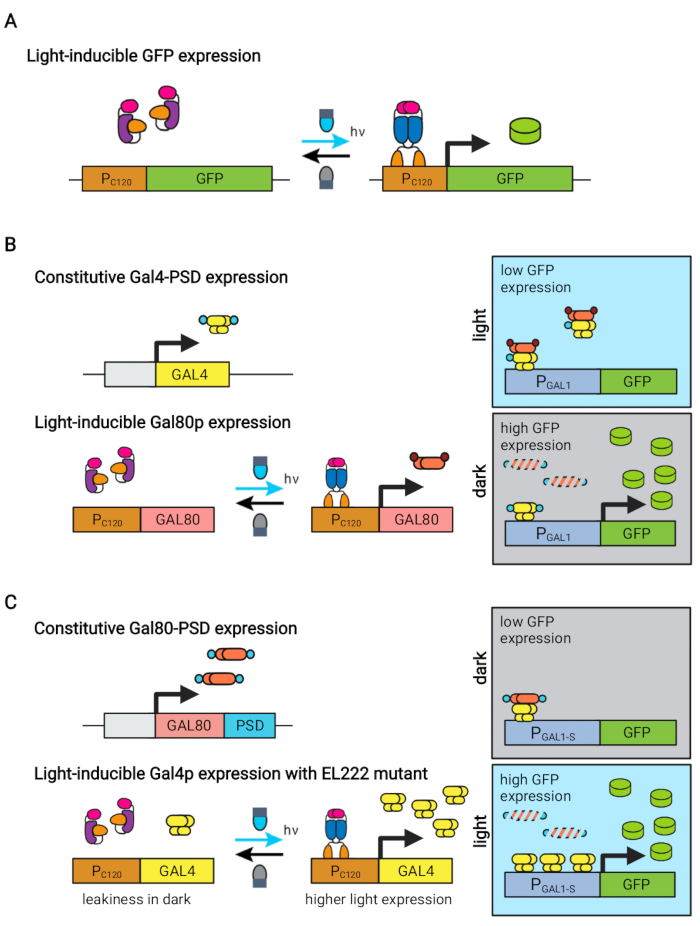
الشكل 1: الدوائر البصرية الجينية للتحكم الديناميكي في S. cerevisiae. تعتمد دوائر OptoEXP و OptoINVRT و OptoAMP على نظام VP16-EL222 الحساس للضوء. (أ) في دائرة OptoEXP ، يؤدي التعرض للضوء الأزرق إلى تغيير توافقي و dimerization من VP16-EL222 ، مما يكشف عن مجال ربط الحمض النووي ويسمح بالنسخ من PC120. وقد عدل هذا الرقم من تشاو وآخرين.7. (B) تسخر دوائر OptoINVRT تنظيمات GAL (المعروضة) أو Q للحث على التعبير في الظلام. في الدوائر القائمة على GAL، يتم التعبير عن VP16-EL222 وGAL4 بشكل تأسيسي، في حين يتم التعبير عن محركات الأقراص PC120 عن مثبط GAL80 (في الدوائر القائمة على Q، يتم استبدال GAL4 وGAL80 ب QF2 و QS، على التوالي، ويتم استخدام مروج اصطناعي يحتوي على QUAS بدلا من مروج GAL). في الضوء ، يمنع Gal80p تنشيط الجين محل الاهتمام من PGAL1. في الظلام ، لا يتم التعبير عن GAL80 ويتحلل بسرعة عن طريق دمجه في مجال degron التأسيسي (المجال البني الصغير) ، والذي يسمح بتنشيط PGAL1 بواسطة Gal4p. وقد عدل هذا الرقم من Zhao et al.5. (C) تستخدم دوائر OptoAMP أيضا VP16-EL222 للتحكم في تنظيمات GAL (المعروضة) أو Q. في هذه الدوائر ، يتم التعبير عن مثبط GAL80 (أو QS) بشكل أساسي ودمجه في degron حساس للصور (مجال أزرق صغير) يضمن القمع المحكم في الظلام. PC120 وتعبير التحكم المتحور VP16-EL222 شديد الحساسية ل GAL4 (أو QF2) مع الضوء ، والذي ينشط بقوة PGAL1 (أو مروج يحتوي على QUAS) في الضوء. يمكن للدوائر المشتقة من GAL استخدام أشكال هندسية من PGAL1 ، مثل PGAL1-M أو PGAL1-S ، والتي زادت من النشاط ، بالإضافة إلى المروجين من النوع البري الذي يتحكم فيه تنظيم GAL (PGAL1 ، PGAL10 ، PGAL2 ، PGAL7). تم تعديل الرقم من Zhao et al.12. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
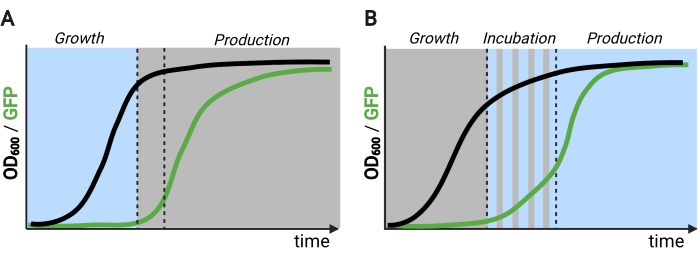
الشكل 2: التخمير ثنائي وثلاثي الطور عبر الزمن. (أ) تتكون عمليات التخمير ثنائية الطور التي تعمل بدوائر مقلوبة من مرحلة نمو مدفوعة بالضوء ومرحلة إنتاج مظلمة. في مرحلة النمو ، تتراكم الكتلة الحيوية مع بقاء مسار الإنتاج مكبوتا. عند الوصول إلى OD600 المطلوب ، يتم نقل الخلايا إلى الظلام لضبط التمثيل الغذائي قبل إعادة تعليقها في وسائط جديدة لمرحلة الإنتاج. (ب) في عملية من ثلاث مراحل ، يتم تحديد مراحل النمو والحضانة والإنتاج من خلال جداول ضوئية فريدة من نوعها ، والتي قد تتكون من فترة نمو مظلمة ، وحضانة نبضية ، ومرحلة إنتاج مضاءة بالكامل. الشكل الذي تم إنشاؤه باستخدام Biorender. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
كما تم تطوير الدوائر البصرية الجينية للتحكم الديناميكي في إنتاج المواد الكيميائية والبروتين في الإشريكية القولونية. تتحكم دوائر OptoLAC في مثبط LacI البكتيري باستخدام دائرة pDawn المستجيبة للضوء ، والتي تستند إلى نظام YF1 / FixJ المكون من مكونين 6 (الشكل 3). على غرار OptoINVRT5 ، تم تصميم دوائر OptoLAC لقمع التعبير الجيني في الضوء وتحفيزه في الظلام. يمكن أن تتطابق مستويات التعبير باستخدام دوائر OptoLAC مع تلك التي تحققت باستخدام تحريض الأيزوبروبيل القياسي β-d-1-thiogalactopyranoside (IPTG) ، وبالتالي الحفاظ على قوة الحث الكيميائي مع توفير إمكانية محسنة للضبط والانعكاس 6. لذلك ، تتيح دوائر OptoLAC التحكم الفعال في البصريات الجينية للهندسة الأيضية في الإشريكية القولونية.
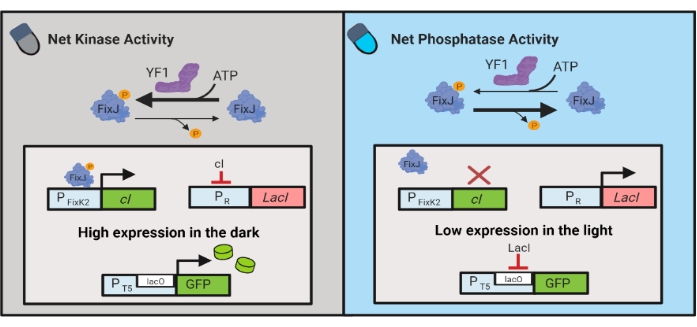
الشكل 3: دوائر OptoLAC للتحكم الديناميكي في الإشريكية القولونية. تقوم دوائر OptoLAC بتكييف نظام pDawn و lac operon لتحقيق التنشيط في الظلام والقمع في الضوء. في الظلام ، يفسفرات YF1 FixJ ، والتي تقوم بعد ذلك بتنشيط مروج PFixK2 للتعبير عن مثبط cI. يمنع مثبط cI التعبير عن مثبط lacI من مروج PR ، مما يسمح بنسخ الجين محل الاهتمام من مروج يحتوي على lacO. على العكس من ذلك ، يقلل الضوء الأزرق من نشاط كيناز صافي YF1 ، مما يعكس فسفرة FixJ وبالتالي تعبير cI ، مما يقلل من التعبير عن lacI ويمنع التعبير من المروج المحتوي على lacO. وقد عدل هذا الرقم من لالواني وآخرين.6. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
نصف هنا البروتوكولات الأساسية للتخمير الذي يتم التحكم فيه بالضوء ل S. cerevisiae و E. coli لإنتاج المواد الكيميائية أو البروتين. بالنسبة لكل من الخميرة والبكتيريا ، نركز أولا على التخمير مع مرحلة نمو مدفوعة بالضوء ومرحلة إنتاج ناجمة عن الظلام ممكنة بواسطة دوائر OptoINVRT و OptoLAC. بعد ذلك ، نصف بروتوكولا للتخمير ثلاثي المراحل (النمو ، الحث ، الإنتاج) الذي يتم التحكم فيه بالضوء والذي يتم تمكينه بواسطة دوائر OptoAMP. علاوة على ذلك ، نصف كيفية توسيع نطاق التخمير الذي يتم التحكم فيه بصريا من الصفائح الدقيقة إلى المفاعلات الحيوية على نطاق المختبر. من خلال هذا البروتوكول ، نهدف إلى توفير دليل كامل وقابل للتكرار بسهولة لإجراء عمليات التخمير التي يتم التحكم فيها بالضوء لإنتاج المواد الكيميائية أو البروتين.
