A optogenética, o controle de processos biológicos com proteínas leves responsivas, oferece uma nova estratégia para controlar dinamicamente as fermentações microbianas para a produção química e proteica1,2. A carga de vias metabólicas projetadas e a toxicidade de alguns intermediários e produtos muitas vezes prejudicam o crescimento celular3. Tais tensões podem levar à má acumulação de biomassa e à redução da produtividade3. Esse desafio pode ser enfrentado dividindo temporalmente as fermentações em uma fase de crescimento e produção, que dedicam recursos metabólicos ao acúmulo de biomassa ou à síntese do produto, respectivamente4. Recentemente, mostramos que a transição do crescimento para a produção nesta fermentação em duas fases pode ser induzida com mudanças nas condições de iluminação5,6,7. A alta sintonia, reversibilidade e ortogonalidade dos insumos leves8 oferecem vantagens únicas às fermentações controladas pela luz que são difíceis ou impossíveis de replicar com indutores químicos usados no controle dinâmico das fermentações convencionais em duas fases4,9,10,11.
A proteína EL222 responsiva de luz azul derivada de Erythrobacter litoralis tem sido usada para desenvolver vários circuitos optogenéticos para engenharia metabólica em Saccharomyces cerevisiae5,7,12,13. El222 contém um domínio sensor de tensão de oxigênio leve (LOV) que sofre uma mudança conformacional sobre a ativação da luz azul (465 nm), que permite que ele se ligue à sua sequência de DNA cognato (C120)13. Fundir EL222 ao domínio de ativação do VP16 viral (VP16-EL222) resulta em um fator de transcrição responsiva de luz azul que pode ativar reversivelmente a expressão genética em S. cerevisiae7 e outros organismos14 do promotor sintético PC120. Vários circuitos baseados no EL222 foram desenvolvidos e utilizados para a produção química em S. cerevisiae, como o sistema OptoEXP ativado pela luz básica7, no qual o gene de interesse é diretamente expresso a partir de PC120 (Figura 1A). No entanto, preocupações com a penetração da luz nas altas densidades celulares tipicamente encontradas na fase de produção de fermentações nos motivaram a desenvolver circuitos invertidos que são induzidos no escuro, como os circuitos OptoINVRT e OptoQ-INVRT (Figura 1B)5,7,13. Estes sistemas aproveitam os regulons galactose (GAL) ou quinic acid (Q) de S. cerevisiae e N. crassa, respectivamente, controlando seus repressores correspondentes (GAL80 e QS) com VP16-EL222, para reprimir a expressão genética na luz e induzi-la fortemente no escuro. A combinação de circuitos OptoEXP e OptoINVRT resulta em controle bidirecional da expressão genética, permitindo fermentações em duas fases nas quais a fase de crescimento é induzida com luz azul, e a fase de produção com escuridão (Figura 2A)5,7.
Usar a luz em vez de escuridão para induzir a expressão genética durante a fase de produção expandiria muito as capacidades de controles optogenéticos, mas também exigiria superar as limitações de penetração de luz das altas densidades celulares tipicamente encontradas nesta fase de fermentação. Para isso, desenvolvemos circuitos, conhecidos como OptoAMP e OptoQ-AMP, que amplificam a resposta transcricional à estimulação da luz azul. Esses circuitos usam mutantes do tipo selvagem ou hipersensíveis do VP16-EL222 para controlar a produção dos ativadores transcricionais Gal4p ou QF2 dos regulons GAL ou Q, respectivamente, alcançando maior sensibilidade e expressão genética mais forte com luz12,13 (Figura 1C). Os circuitos optoAMP podem alcançar indução de luz completa e homogênea em bioreatores de 5 L a uma densidade óptica (medida a 600 nm; OD600) valores de pelo menos 40 com apenas ~0,35% de iluminação (5% de dose leve em apenas ~7% da superfície a granel). Isso demonstra um maior grau de sensibilidade em comparação com o OptoEXP, que requer cerca de 100% de iluminação12. A capacidade de induzir efetivamente a expressão genética com luz em altas densidades celulares abre novas oportunidades para o controle dinâmico das fermentações. Isso inclui fermentações operacionais em mais de duas fases temporais, como fermentações trifásicas, nas quais as fases de crescimento, indução e produção são estabelecidas com cronogramas de luz únicos para otimizar a produção química (Figura 2B)12.
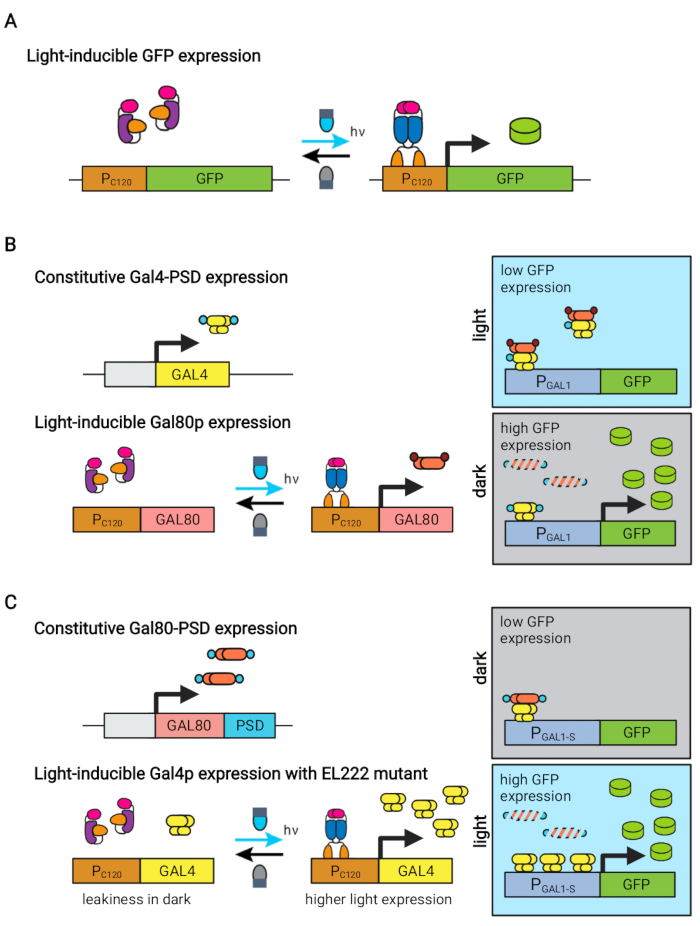
Figura 1: Circuitos optogenéticos para controle dinâmico de S. cerevisiae. Os circuitos OptoEXP, OptoINVRT e OptoAMP são baseados no sistema VP16-EL222 sensível à luz. (A) No circuito OptoEXP, a exposição à luz azul causa uma alteração conformacional e a dimerização do VP16-EL222, que expõe um domínio de vinculação de DNA e permite a transcrição do PC120. O número foi modificado de Zhao et al.7. (B) Os circuitos OptoINVRT aproveitam os regulons GAL (mostrado) ou Q para induzir a expressão no escuro. Nos circuitos baseados em GAL, VP16-EL222 e GAL4 são expressos constitutivamente, enquanto a expressão de drives PC120 do repressor GAL80 (em circuitos baseados em Q, GAL4 e GAL80 são substituídas por QF2 e QS, respectivamente, e um promotor sintético contendo QUAS é usado em vez de um promotor gal). À luz, Gal80p impede a ativação do gene de interesse do PGAL1. No escuro, GAL80 não é expresso e rapidamente degradado, fundindo-o a um domínio degron constitutivo (pequeno domínio marrom), que permite a ativação de PGAL1 por Gal4p. O número foi modificado de Zhao et al.5. (C) Os circuitos OptoAMP também usam VP16-EL222 para controlar os regulons GAL (mostrado) ou Q. Nestes circuitos, o repressor GAL80 (ou QS) é expresso e fundido constitutivamente e fundido a um degron sensível a fotos (pequeno domínio azul) garantindo uma repressão apertada no escuro. PC120 e uma expressão de controle mutante hipersensível VP16-EL222 de GAL4 (ou QF2) com luz, que ativa fortemente O PGAL1 (ou um promotor contendo QUAS) na luz. Os circuitos derivados de GAL podem usar formas projetadas de PGAL1, como PGAL1-M ou PGAL1-S, que aumentaram a atividade, bem como promotores do tipo selvagem controlados pelo regulon GAL (PGAL1, PGAL10, PGAL2, PGAL7). A figura foi modificada de Zhao et al.12. Clique aqui para ver uma versão maior desta figura.
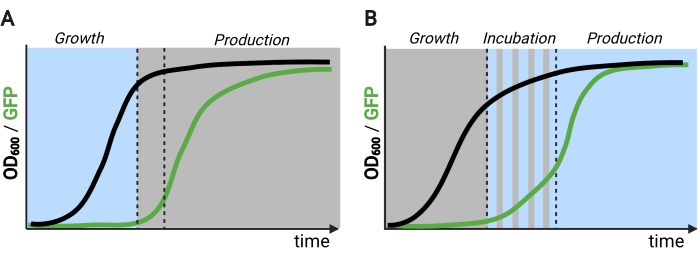
Figura 2: Fermentações de duas e três fases através do tempo. (A) Fermentações em duas fases operadas com circuitos invertidos consistem em uma fase de crescimento orientada pela luz e uma fase de produção escura. Na fase de crescimento, a biomassa se acumula à medida que a via de produção permanece reprimida. Ao atingir o OD600 desejado, as células são deslocadas para o escuro para ajustar metabolicamente antes de serem resuspendidas em novas mídias para a fase de produção. (B) Em um processo trifásica, as fases de crescimento, incubação e produção são definidas por cronogramas de luz únicos, que podem consistir em um período de crescimento escuro, incubação pulsada e fase de produção totalmente iluminada. Figura criada com Biorender. Clique aqui para ver uma versão maior desta figura.
Circuitos optogenéticos também foram desenvolvidos para controle dinâmico da produção química e proteica em E. coli. Os circuitos optoLAC controlam o repressor LacI bacteriano usando o circuito pDawn responsivo à luz, que é baseado no sistema de dois componentes YF1/FixJ6 (Figura 3). Semelhante ao OptoINVRT5, os circuitos OptoLAC são projetados para reprimir a expressão genética na luz e induzi-la no escuro. Os níveis de expressão usando circuitos OptoLAC podem corresponder ou exceder aqueles alcançados com indução isopropílico padrão β-d-1-thiogalactopyranoside (IPTG), mantendo assim a força da indução química ao mesmo tempo que oferece maior sintonia e reversibilidade6. Portanto, os circuitos OptoLAC permitem um controle optogenético eficaz para a engenharia metabólica em E. coli.
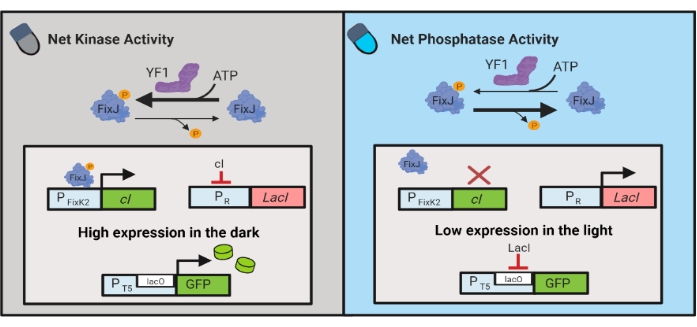
Figura 3: Circuitos OptoLAC para controle dinâmico de E. coli. Os circuitos OptoLAC adaptam o sistema pDawn e operon de lac para alcançar ativação no escuro e repressão à luz. No escuro, o YF1 fosforila fixJ, que então ativa o promotor PFixK2 para expressar o repressor cI . O repressor de IC impede a expressão do repressor lacI do promotor de PR , que permite a transcrição do gene de interesse de um promotor contendo lacO. Por outro lado, a luz azul reduz a atividade de quinase líquida YF1, invertendo a fosforilação fixJ e, portanto, a expressão cI , que deprime a expressão do lacI e impede a expressão do promotor contendo lacO. O número foi modificado de Lalwani et al.6. Clique aqui para ver uma versão maior desta figura.
Descrevemos aqui os protocolos básicos para fermentações controladas pela luz de S. cerevisiae e E. coli para produção química ou proteica. Para a levedura e as bactérias, primeiro focamos em fermentações com uma fase de crescimento impulsionada pela luz e uma fase de produção induzida pela escuridão habilitada pelos circuitos OptoINVRT e OptoLAC. Posteriormente, descrevemos um protocolo para uma fermentação trifásica (crescimento, indução, produção) controlada por luz habilitada pelos circuitos OptoAMP. Além disso, descrevemos como escalar fermentações optogeneticamente controladas de microplacos a bioreatores em escala de laboratório. Com este protocolo, pretendemos fornecer um guia completo e facilmente reprodutível para a realização de fermentações controladas por luz para produção química ou proteica.
