אופטוגנטיקה, השליטה בתהליכים ביולוגיים עם חלבונים מגיבים לאור, מציעה אסטרטגיה חדשה לשליטה דינמית בתסיסות מיקרוביות לייצור כימי וחלבון 1,2. הנטל של מסלולים מטבוליים מהונדסים ואת הרעילות של כמה מתווכים ומוצרים לעתים קרובות פוגע צמיחת התא3. לחצים כאלה יכולים להוביל הצטברות ביומסה לקויה ופרודוקטיביות מופחתת3. אתגר זה ניתן לטפל על ידי חלוקה זמנית תסיסה לשלב צמיחה וייצור, אשר מקדישים משאבים מטבוליים הצטברות ביומסה או סינתזת מוצר בהתאמה4. לאחרונה הראינו כי המעבר מצמיחה לייצור בתסיסה דו-שלבית זו יכול להיות מושרה עם שינויים בתנאי תאורה5,6,7. הטונה הגבוהה, הפיכות והאורתוגונליות של תשומות אור8 מציעות יתרונות ייחודיים לתסיסות מבוקרות אור שקשה או בלתי אפשרי לשכפל עם ממריצים כימיים המשמשים לשליטה דינמית בתסיסות דו-פאזיות קונבנציונליות4,9,10,111.
חלבון EL222 מגיב אור כחול נגזר ליטורליס אריתרובקטריה שימש לפיתוח מספר מעגלים אופטוגנטיים להנדסה מטבולית ב Saccharomyces cerevisiae5,7,12,13. EL222 מכיל תחום חיישן מתח אור-חמצן (LOV) שעובר שינוי קונפורמציה עם הפעלת אור כחול (465 ננומטר), המאפשר לו להיקשר לרצף הדנ”א ההכרה שלו (C120)13. היתוך EL222 לתחום ההפעלה הנגיפי VP16 (VP16-EL222) מביא לגורם שעתוק מגיב לאור כחול שיכול להפעיל באופן הפיך ביטוי גנים ב- S. cerevisiae7 ובאורגניזמים אחרים14 מהמקדם הסינתטי PC120. מספר מעגלים המבוססים על EL222 פותחו ושימשו לייצור כימי ב- S. cerevisiae, כגון מערכת OptoEXP הבסיסית המופעלת באמצעות אור7, שבה גן העניין בא לידי ביטוי ישירות מ- PC120 (איור 1A). עם זאת, החששות מפני חדירת אור בצפיפות התאים הגבוהה שבדרך כלל נתקלים בהם בשלב הייצור של התסיסות, הניעה אותנו לפתח מעגלים הפוכים המושרים בחושך, כגון מעגלי OptoINVRT ו- OptoQ-INVRT (איור 1B)5,7,13. מערכות אלה לרתום את הגלקטוז (GAL) או חומצה קווינית (Q) regulons מ S. cerevisiae ו N. crassa, בהתאמה, שליטה על הדיכויים המקבילים שלהם (GAL80 ו QS) עם VP16-EL222, כדי להדחיק ביטוי גנים באור ולגרום לו חזק בחושך. שילוב מעגלי OptoEXP ו-OptoINVRT מביא לשליטה דו-כיוונית בביטוי הגנים, ומאפשר תסיסה דו-שלבית שבה שלב הצמיחה מושרה באור כחול, ושלב הייצור עם חושך (איור 2A)5,7.
שימוש באור במקום בחושך כדי לגרום לביטוי גנים במהלך שלב הייצור ירחיב מאוד את היכולות של בקרות אופטוגנטיות, אך גם ידרוש התגברות על מגבלות חדירת האור של צפיפות התאים הגבוהה שבדרך כלל נתקלת בה בשלב זה של תסיסה. לשם כך, פיתחנו מעגלים, המכונים OptoAMP ו- OptoQ-AMP, המגבירים את תגובת התמלול לגירוי אור כחול. מעגלים אלה משתמשים במוטנטים פראיים או רגישים יתר על המידה של VP16-EL222 כדי לשלוט בייצור של מפעילי התמלול Gal4p או QF2 של regulons GAL או Q, בהתאמה, השגת רגישות משופרת וביטוי גנים חזק יותר עם אור 12,13 (איור 1C). מעגלי OptoAMP יכולים להשיג אינדוקציה מלאה והומוגנית של אור ב-5 L bioreactors בצפיפות אופטית (נמדדת ב-600 ננומטר; OD600) ערכים של לפחות 40 עם רק ~0.35% של תאורה (5% מינון אור רק ~ 7% של פני השטח בתפזורת). זה מדגים רמה גבוהה יותר של רגישות לעומת OptoEXP, אשר דורש קרוב 100% תאורה12. היכולת לגרום ביעילות לביטוי גנים עם אור בצפיפות תאים גבוהה פותחת הזדמנויות חדשות לשליטה דינמית בתסיסות. זה כולל תסיסה תפעולית ביותר משני שלבים זמניים, כגון תסיסה תלת-פאזית, שבה שלבי צמיחה, אינדוקציה וייצור נקבעים עם לוחות זמנים ייחודיים של אור כדי לייעל את הייצור הכימי (איור 2B)12.
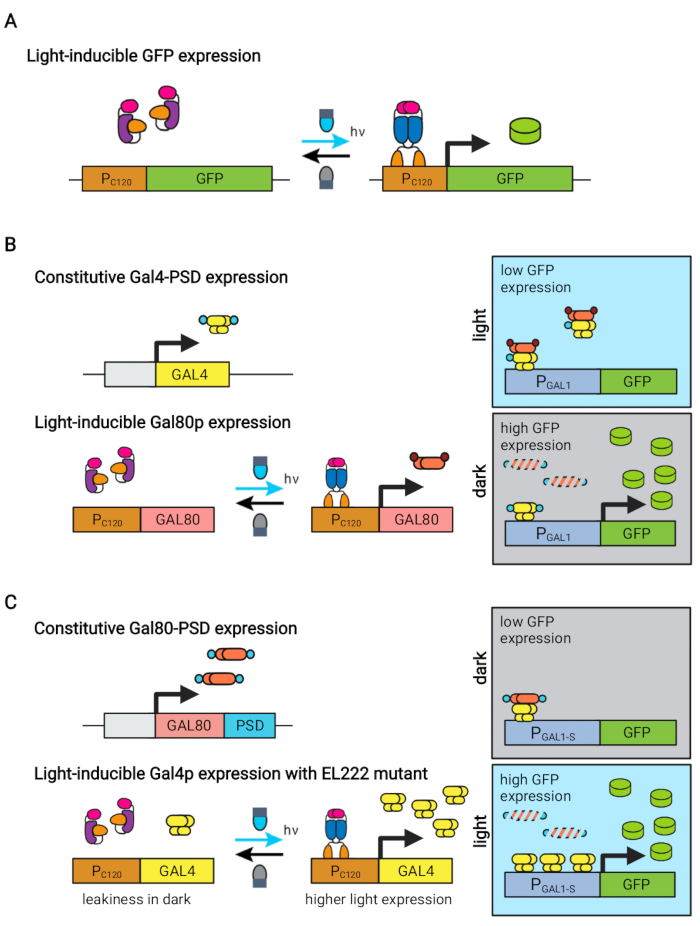
איור 1: מעגלים אופטוגנטיים לשליטה דינמית ב-S. cerevisiae. מעגלי OptoEXP, OptoINVRT ו-OptoAMP מבוססים על מערכת VP16-EL222 הרגישה לאור. (א) במעגל OptoEXP, חשיפה לאור כחול גורמת לשינוי קונפורמציה ודימום של VP16-EL222, החושף תחום מחייב DNA ומאפשר שעתוק מ- PC120. הנתון שונה מז’או ואח’ 7. (ב) מעגלי OptoINVRT רותמים את התגמולונים GAL (המוצגים) או Q כדי לגרום לביטוי בחושך. במעגלים מבוססי GAL, VP16-EL222 ו- GAL4 באים לידי ביטוי באופן מכונן, בעוד PC120 מניע ביטוי של מדכא GAL80 (במעגלים מבוססי Q, GAL4 ו- GAL80 מוחלפים על ידי QF2 ו- QS, בהתאמה, ומקדם סינתטי המכיל QUAS משמש במקום מקדם GAL). לאור זאת, Gal80p מונע הפעלה של הגן של עניין מ PGAL1. בחושך, GAL80 אינו בא לידי ביטוי ומושפל במהירות על ידי היתוך אותו לתחום degron מכונן (תחום חום קטן), המאפשר הפעלה של PGAL1 על ידי Gal4p. הנתון שונה מז’או ואח’ 5. (ג) מעגלי OptoAMP משתמשים גם ב-VP16-EL222 כדי לשלוט ברגולונים GAL (מוצגים) או Q. במעגלים אלה, מדכא GAL80 (או QS) מבוטא באופן מכונן ומותך לדגרון רגיש לצילום (תחום כחול קטן) המבטיח דיכוי הדוק בחושך. PC120 וביטוי בקרת מוטציה VP16-EL222 רגיש של GAL4 (או QF2) עם אור, אשר מפעיל בעוצמה PGAL1 (או מקדם המכיל QUAS) באור. מעגלים שמקורם ב-GAL יכולים להשתמש בצורות מהונדסות של PGAL1, כגון PGAL1-M או PGAL1-S, אשר הגבירו את הפעילות, כמו גם מקדמים מסוג בר הנשלטים על ידי ה-GAL regulon (PGAL1, PGAL10, PGAL2, PGAL7). האיור שונה מ- Zhao et al.12. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
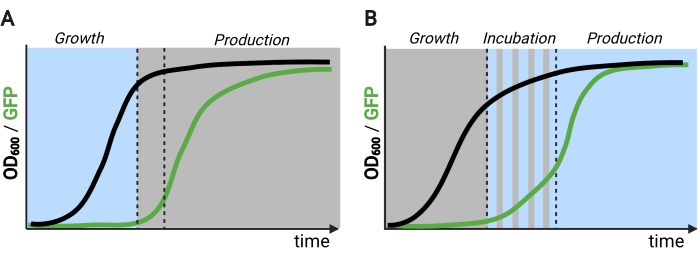
איור 2: תסיסה דו-שלבית ותלת-פאזית לאורך זמן. (A) תסיסה דו-פאזית המופעלת עם מעגלים הפוכים מורכבת משלב צמיחה מונחה אור ושלב ייצור כהה. בשלב הצמיחה, ביומסה מצטברת כאשר מסלול הייצור נשאר מודחק. עם ההגעה OD600 הרצוי, תאים מועברים אל החושך כדי להתאים מטבולית לפני להיות resuspended במדיה טרייה עבור שלב הייצור. (ב) בתהליך תלת-שלבי, שלבי הצמיחה, הדגירה והייצור מוגדרים על ידי לוחות זמנים ייחודיים של אור, שעשויים להיות מורכבים מתקופה של צמיחה חשוכה, דגירה פועמת ושלב ייצור מואר לחלוטין. דמות שנוצרה עם ביורנדר. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
מעגלים אופטוגנטיים פותחו גם לשליטה דינמית בייצור כימי וחלבון ב– E. coli. מעגלי OptoLAC שולטים במדכא LacI החיידקי באמצעות מעגל pDawn בעל תגובת האור, המבוסס על מערכת שני הרכיבים YF1/FixJ6 (איור 3). בדומה ל- OptoINVRT5, מעגלי OptoLAC נועדו להדחיק ביטוי גנים באור ולגרום לו בחושך. רמות הביטוי באמצעות מעגלי OptoLAC יכולות להתאים או לחרוג מאלה שהושגו עם אינדוקציה סטנדרטית של איזופרופיל β-d-1-thiogalactopyranoside (IPTG), ובכך לשמור על כוח האינדוקציה הכימית תוך מתן טונה והפכה משופרות6. לכן, מעגלי OptoLAC מאפשרים שליטה אופטוגנטית יעילה להנדסה מטבולית ב– E. coli.
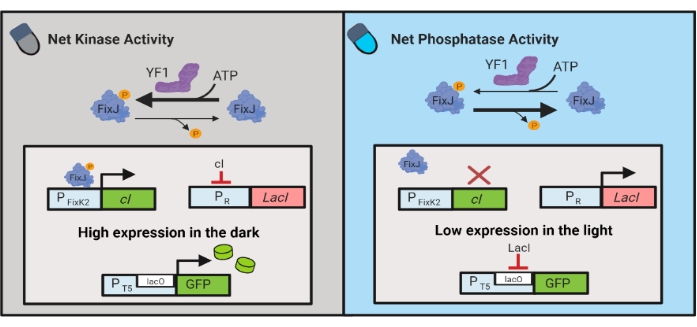
איור 3: מעגלי OptoLAC לשליטה דינמית ב-E. coli. מעגלי OptoLAC מתאימים את מערכת ה-pDawn ואת אופרון הלק כדי להשיג הפעלה בחושך ודיכוי באור. בחושך, YF1 זרחן FixJ, אשר לאחר מכן מפעיל את מקדם PFixK2 להביע את מדכא cI . מדכא ה – cI מונע ביטוי של מדחיק lacI ממקדם ה- PR , המאפשר שעתוק של גן העניין ממקדם המכיל לאקו. לעומת זאת, אור כחול מפחית את פעילות קינאז נטו YF1, היפוך זרחן FixJ ובכך ביטוי cI , אשר מפחית את הביטוי של lacI ומונע ביטוי מהמקדם המכיל לאקו. הנתון שונה מ Lalwani et al.6. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
אנו מתארים כאן את הפרוטוקולים הבסיסיים לתסיסות מבוקרות אור של S. cerevisiae ו – E. coli לייצור כימי או חלבון. עבור שמרים וחיידקים כאחד, אנו מתמקדים תחילה בתסיסות עם שלב צמיחה מונחה אור ושלב ייצור המושרה בחושך המופעל על ידי מעגלי OptoINVRT ו- OptoLAC. לאחר מכן, אנו מתארים פרוטוקול לתסיסה תלת-פאזית (צמיחה, אינדוקציה, ייצור) מבוקרת אור המופעלת על ידי מעגלי OptoAMP. יתר על כן, אנו מתארים כיצד להרחיב תסיסה מבוקרת אופטוגנטית ממיקרו-לוחות ל-bioreactors בקנה מידה של מעבדה. עם פרוטוקול זה, אנו שואפים לספק מדריך מלא וקל לשחזור לביצוע תסיסה מבוקרת אור לייצור כימי או חלבון.
