Optogenetik, kontrol af biologiske processer med lysresponsive proteiner, tilbyder en ny strategi til dynamisk kontrol af mikrobielle fermentementer til kemisk og proteinproduktion1,2. Byrden ved konstruerede metaboliske veje og toksiciteten af visse mellemprodukter og produkter forringer ofte cellevæksten3. Sådanne belastninger kan føre til dårlig ophobning af biomasse og nedsat produktivitet3. Denne udfordring kan løses ved tidsmæssigt at opdele fermenteringen i en vækst- og produktionsfase, som afsætter metaboliske ressourcer til henholdsvis biomasseakkumulering eller produktsyntese4. Vi viste for nylig, at overgangen fra vækst til produktion i denne tofasede gæring kan induceres med ændringer i belysningsforholdene5,6,7. Lysindgangens høje tunbarhed, reversibilitet og ortogonalitet8 giver unikke fordele ved lysstyrede fermenteringener, der er vanskelige eller umulige at replikere med kemiske induktorer, der anvendes til dynamisk kontrol af konventionelle tofasede fermenteringener4,9,10,11.
Det blå lys responsive EL222 protein afledt af Erythrobacter litoralis er blevet brugt til at udvikle flere optogenetiske kredsløb til metabolisk teknik i Saccharomyces cerevisiae5,7,12,13. EL222 indeholder et lys-ilt-spændingssensor (LOV) domæne, der gennemgår et konformationsskift ved aktivering af blåt lys (465 nm), som gør det muligt at binde sig til sin beslægtede DNA-sekvens (C120)13. Sammensmeltning af EL222 til det virale VP16-aktiveringsdomæne (VP16-EL222) resulterer i en blålysresponsiv transkriptionsfaktor, der reversibelt kan aktivere genekspression i S. cerevisiae7 og andre organismer14 fra den syntetiske promotor PC120. Flere kredsløb baseret på EL222 er blevet udviklet og anvendt til kemisk produktion i S. cerevisiae, såsom det grundlæggende lysaktiverede OptoEXP-system7, hvor genet af interesse udtrykkes direkte fra PC120 (figur 1A). Bekymringer om lysindtrængning ved de høje celletætheder, der typisk opstår i produktionsfasen af fermenteringer, motiverede os imidlertid til at udvikle omvendte kredsløb, der induceres i mørket, såsom OptoINVRT- og OptoQ-INVRT-kredsløbene (figur 1B) 5,7,13. Disse systemer udnytter galactose (GAL) eller dikinsyre (Q) reguloner fra henholdsvis S. cerevisiae og N. crassa, der styrer deres tilsvarende repressorer (GAL80 og QS) med VP16-EL222, for at undertrykke genekspression i lyset og stærkt inducere det i mørket. Kombination af OptoEXP- og OptoINVRT-kredsløb resulterer i tovejskontrol af genekspression, hvilket muliggør tofasede fermenteringer, hvor vækstfasen induceres med blåt lys, og produktionsfasen med mørke (figur 2A)5,7.
Brug af lys i stedet for mørke til at fremkalde genekspression i produktionsfasen ville i høj grad udvide mulighederne for optogenetiske kontroller, men ville også kræve at overvinde lysindtrængningsbegrænsningerne for de høje celletætheder, der typisk opstår i denne fase af gæringen. Til dette formål har vi udviklet kredsløb, kendt som OptoAMP og OptoQ-AMP, der forstærker transkriptionsresponsen på stimulering af blåt lys. Disse kredsløb bruger vildtype eller overfølsomme mutanter af VP16-EL222 til at kontrollere produktionen af transkriptionsaktivatorerne Gal4p eller QF2 af henholdsvis GAL- eller Q-regulonerne, hvilket opnår øget følsomhed og stærkere genekspression med lys12,13 (figur 1C). OptoAMP-kredsløb kan opnå fuldstændig og homogen lysinduktion i 5 L bioreaktorer ved en optisk densitet (målt ved 600 nm; OD600) værdier på mindst 40 med kun ~0,35% af belysningen (5% lysdosis på kun ~7% af bulkoverfladen). Dette viser en højere grad af følsomhed sammenlignet med OptoEXP, som kræver tæt på 100 % belysning12. Evnen til effektivt at inducere genekspression med lys ved høje celletætheder åbner nye muligheder for dynamisk kontrol af fermenteringer. Dette omfatter driftsgæringer i mere end to tidsmæssige faser, såsom trefasede fermentementer, hvor vækst-, induktions- og produktionsfaser etableres med unikke lysplaner for at optimere kemisk produktion (figur 2B)12.
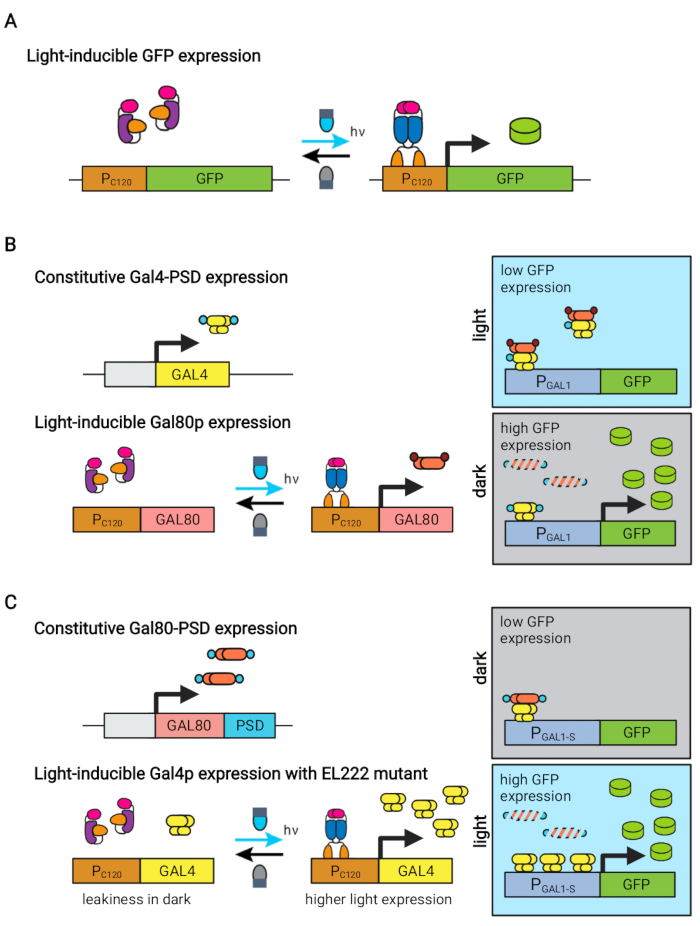
Figur 1: Optogenetiske kredsløb til dynamisk styring af S. cerevisiae. OptoEXP-, OptoINVRT- og OptoAMP-kredsløbene er baseret på det lysfølsomme VP16-EL222-system. (A) I OptoEXP-kredsløbet forårsager eksponering for blåt lys en konformationsændring og dimerisering af VP16-EL222, som udsætter et DNA-bindende domæne og muliggør transkription fra PC120. Figuren er blevet modificeret fra Zhao et al.7. (B) OptoINVRT-kredsløb udnytter GAL-regulonerne (vist) eller Q-regulonerne til at fremkalde udtryk i mørket. I GAL-baserede kredsløb udtrykkes VP16-EL222 og GAL4 konstitutivt, mens PC120-drevudtryk for GAL80-repressoren (i Q-baserede kredsløb erstattes GAL4 og GAL80 af henholdsvis QF2 og QS, og en syntetisk QUAS-holdig promotor anvendes i stedet for en GAL-promotor). I lyset forhindrer Gal80p aktivering af genet af interesse fra PGAL1. I mørket udtrykkes GAL80 ikke og nedbrydes hurtigt ved at smelte det sammen til et konstitutivt degrondomæne (lille brunt domæne), hvilket gør det muligt at aktivere PGAL1 af Gal4p. Figuren er blevet modificeret fra Zhao et al.5. (C) OptoAMP-kredsløb bruger også VP16-EL222 til at styre GAL(vist) eller Q-regulonerne. I disse kredsløb udtrykkes GAL80-repressoren (eller QS) konstitutivt og smeltes sammen til en fotofølsom degron (lille blåt domæne), der sikrer tæt undertrykkelse i mørket. PC120 og en overfølsom VP16-EL222 mutant kontrolekspression af GAL4 (eller QF2) med lys, som stærkt aktiverer PGAL1 (eller en QUAS-holdig promotor) i lyset. GAL-afledte kredsløb kan bruge konstruerede former for PGAL1, såsom PGAL1-M eller PGAL1-S, som har øget aktivitet, samt vildtypepromotorer kontrolleret af GAL-regulon (PGAL1, PGAL10, PGAL2, PGAL7). Figuren er ændret fra Zhao et al.12. Klik her for at se en større version af denne figur.
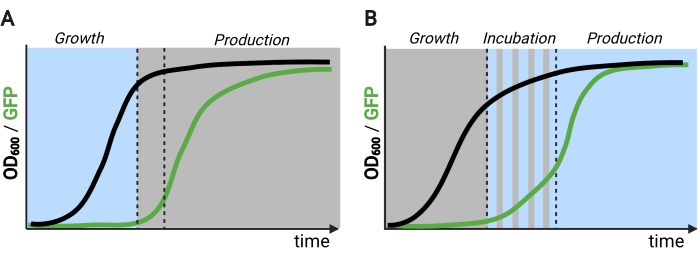
Figur 2: To- og trefasede fermentementer gennem tiden. (A) Tofasede fermentementer, der drives med inverterede kredsløb, består af en lysdrevet vækstfase og en mørk produktionsfase. I vækstfasen akkumuleres biomasse, da produktionsvejen forbliver undertrykt. Efter at have nået den ønskede OD600, skiftes cellerne til mørket for metabolisk justering, inden de resuspendes i friske medier til produktionsfasen. (B) I en trefaset proces defineres vækst-, inkubations- og produktionsfaserne af unikke lysskemaer, som kan bestå af en mørk vækstperiode, pulserende inkubation og fuldt belyst produktionsfase. Figur oprettet med Biorender. Klik her for at se en større version af denne figur.
Optogenetiske kredsløb er også udviklet til dynamisk styring af kemisk og proteinproduktion i E. coli. OptoLAC-kredsløb styrer den bakterielle LacI-repressor ved hjælp af det lysresponsive pDawn-kredsløb, som er baseret på YF1/FixJ-tokomponentsystemet6 (figur 3). I lighed med OptoINVRT5 er OptoLAC-kredsløb designet til at undertrykke genekspression i lyset og inducere det i mørket. Ekspressionsniveauer ved hjælp af OptoLAC-kredsløb kan matche eller overstige dem, der opnås med standard isopropyl-β-d-1-thiogalactopyranosid (IPTG) induktion, hvilket opretholder styrken af kemisk induktion, samtidig med at den giver forbedret tunbarhed og reversibilitet6. Derfor muliggør OptoLAC-kredsløb effektiv optogenetisk kontrol til metabolisk teknik i E. coli.
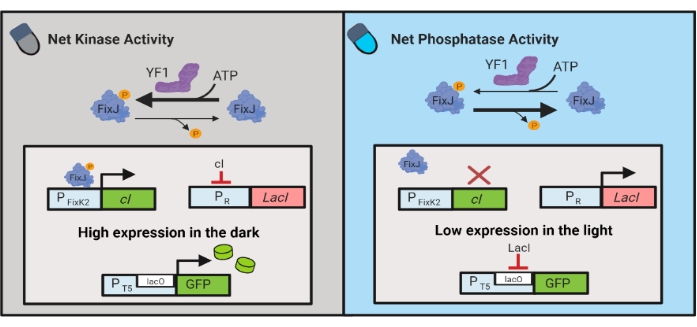
Figur 3: OptoLAC-kredsløb til dynamisk styring af E. coli. OptoLAC-kredsløbene tilpasser pDawn-systemet og lac operon for at opnå aktivering i mørket og undertrykkelse i lyset. I mørket fosforylerer YF1 FixJ, som derefter aktiverer PFixK2-promotoren for at udtrykke cI-repressoren. CI-repressoren forhindrer ekspression af lacI-repressoren fra PR-promotoren, hvilket tillader transkription af genet af interesse fra en lacO-holdige promotor. Omvendt reducerer blåt lys YF1-netkinaseaktiviteten, vender FixJ-fosforylering og dermed cI-ekspression, hvilket aflaster ekspression af lacI og forhindrer ekspression fra den lacO-holdige promotor. Figuren er modificeret fra Lalwani et al.6. Klik her for at se en større version af denne figur.
Vi beskriver her de grundlæggende protokoller for lyskontrollerede fermenteringen af S. cerevisiae og E. coli til kemisk eller proteinproduktion. For både gær og bakterier fokuserer vi først på fermentementer med en lysdrevet vækstfase og en mørkeinduceret produktionsfase muliggjort af OptoINVRT- og OptoLAC-kredsløb. Derefter beskriver vi en protokol for en trefaset (vækst, induktion, produktion) lysstyret gæring aktiveret af OptoAMP-kredsløb. Desuden beskriver vi, hvordan man opskalerer optogenetisk kontrollerede fermentementer fra mikroplader til laboratorieskala bioreaktorer. Med denne protokol sigter vi mod at levere en komplet og let reproducerbar vejledning til udførelse af lyskontrollerede fermenteringener til kemisk eller proteinproduktion.
