L’optogenetica, il controllo dei processi biologici con proteine sensibili alla luce, offre una nuova strategia per controllare dinamicamente le fermentazioni microbiche per la produzione chimica e proteica1,2. L’onere delle vie metaboliche ingegnerizzate e la tossicità di alcuni intermedi e prodotti spesso compromettono la crescita cellulare3. Tali sollecitazioni possono portare a uno scarso accumulo di biomassa e a una ridotta produttività3. Questa sfida può essere affrontata dividendo temporalmente le fermentazioni in una fase di crescita e produzione, che dedicano risorse metaboliche rispettivamente all’accumulo di biomassa o alla sintesi del prodotto4. Recentemente abbiamo dimostrato che il passaggio dalla crescita alla produzione in questa fermentazione bifase può essere indotto con cambiamenti nelle condizioni di illuminazione5,6,7. L’elevata sintonizzazione, reversibilità e ortogonalità degli ingressi luminosi8 offre vantaggi unici alle fermentazioni controllate dalla luce che sono difficili o impossibili da replicare con induttori chimici utilizzati nel controllo dinamico delle fermentazioni bifase convenzionali4,9,10,11.
La proteina EL222 sensibile alla luce blu derivata da Erythrobacter litoralis è stata utilizzata per sviluppare diversi circuiti optogenetici per l’ingegneria metabolica in Saccharomyces cerevisiae5,7,12,13. EL222 contiene un dominio LOV (Light-Oxygen-Voltage Sensor) che subisce uno spostamento conformazionale all’attivazione della luce blu (465 nm), che gli consente di legarsi alla sua sequenza di DNA affine (C120)13. La fusione di EL222 con il dominio di attivazione virale VP16 (VP16-EL222) si traduce in un fattore di trascrizione sensibile alla luce blu che può attivare in modo reversibile l’espressione genica in S. cerevisiae7 e altri organismi14 dal promotore sintetico PC120. Diversi circuiti basati su EL222 sono stati sviluppati e utilizzati per la produzione chimica in S. cerevisiae, come il sistema optoEXP di base attivato dalla luce7, in cui il gene di interesse è espresso direttamente da PC120 (Figura 1A). Tuttavia, le preoccupazioni di penetrazione della luce alle alte densità cellulari tipicamente incontrate nella fase di produzione delle fermentazioni ci hanno motivato a sviluppare circuiti invertiti indotti al buio, come i circuiti OptoINVRT e OptoQ-INVRT (Figura 1B)5,7,13. Questi sistemi sfruttano i regolaloni di galattosio (GAL) o acido chinico (Q) di S. cerevisiae e N. crassa, rispettivamente, controllando i loro corrispondenti repressori (GAL80 e QS) con VP16-EL222, per reprimere l’espressione genica alla luce e indurla fortemente al buio. La combinazione dei circuiti OptoEXP e OptoINVRT si traduce in un controllo bidirezionale dell’espressione genica, consentendo fermentazioni bifase in cui la fase di crescita è indotta con luce blu e la fase di produzione con oscurità (Figura 2A)5,7.
L’uso della luce al posto dell’oscurità per indurre l’espressione genica durante la fase di produzione espanderebbe notevolmente le capacità dei controlli optogenetici, ma richiederebbe anche il superamento dei limiti di penetrazione della luce delle alte densità cellulari tipicamente incontrate in questa fase di fermentazione. A tal fine, abbiamo sviluppato circuiti, noti come OptoAMP e OptoQ-AMP, che amplificano la risposta trascrizionale alla stimolazione della luce blu. Questi circuiti utilizzano mutanti wild-type o ipersensibili di VP16-EL222 per controllare la produzione degli attivatori trascrizionali Gal4p o QF2 dei reguloni GAL o Q, rispettivamente, ottenendo una maggiore sensibilità e una maggiore espressione genica con la luce12,13 (Figura 1C). I circuiti OptoAMP possono ottenere un’induzione luminosa completa e omogenea in bioreattori da 5 L a densità ottica (misurata a 600 nm; OD600) valori di almeno 40 con solo ~0,35% di illuminazione (5% di dose di luce solo su ~7% della superficie di massa). Ciò dimostra un grado di sensibilità più elevato rispetto a OptoEXP, che richiede un’illuminazione vicina al 100%12. La capacità di indurre efficacemente l’espressione genica con la luce ad alte densità cellulari apre nuove opportunità per il controllo dinamico delle fermentazioni. Ciò include fermentazioni operative in più di due fasi temporali, come le fermentazioni trifase, in cui le fasi di crescita, induzione e produzione sono stabilite con programmi leggeri unici per ottimizzare la produzione chimica (Figura 2B)12.
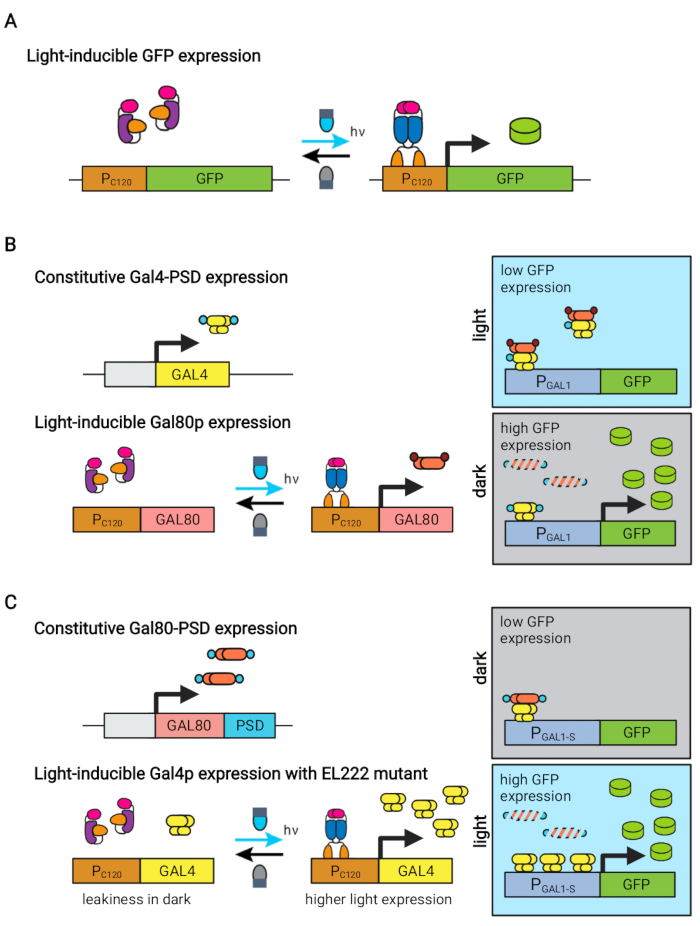
Figura 1: Circuiti optogenetici per il controllo dinamico di S. cerevisiae. I circuiti OptoEXP, OptoINVRT e OptoAMP si basano sul sistema VP16-EL222 sensibile alla luce. (A) Nel circuito OptoEXP, l’esposizione alla luce blu provoca un cambiamento conformazionale e la dimerizzazione di VP16-EL222, che espone un dominio legante il DNA e consente la trascrizione da PC120. La figura è stata modificata da Zhao et al.7. (B) I circuiti OptoINVRT sfruttano i regolagni GAL (mostrati) o Q per indurre l’espressione al buio. Nei circuiti basati su GAL, VP16-EL222 e GAL4 sono espressi costitutivamente, mentre PC120 guida l’espressione del repressore GAL80 (nei circuiti basati su Q, GAL4 e GAL80 sono sostituiti rispettivamente da QF2 e QS e viene utilizzato un promotore sintetico contenente QUAS al posto di un promotore GAL). Alla luce, Gal80p impedisce l’attivazione del gene di interesse da PGAL1. Al buio, GAL80 non è espresso e rapidamente degradato fondendolo in un dominio costitutivo di degron (piccolo dominio marrone), che consente l’attivazione di PGAL1 da parte di Gal4p. La figura è stata modificata da Zhao et al.5. (C) I circuiti OptoAMP utilizzano anche VP16-EL222 per controllare i reguloni GAL (mostrato) o Q. In questi circuiti, il repressore GAL80 (o QS) è costitutivamente espresso e fuso in un degron fotosensibile (piccolo dominio blu) che garantisce una stretta repressione al buio. PC120 e un’espressione di controllo mutante ipersensibile VP16-EL222 di GAL4 (o QF2) con la luce, che attiva fortemente PGAL1 (o un promotore contenente QUAS) nella luce. I circuiti derivati da GAL possono utilizzare forme ingegnerizzate di PGAL1, come PGAL1-M o PGAL1-S, che hanno aumentato l’attività, così come promotori wild-type controllati dal regulon GAL (PGAL1, PGAL10, PGAL2, PGAL7). La figura è stata modificata da Zhao et al.12. Fare clic qui per visualizzare una versione più grande di questa figura.
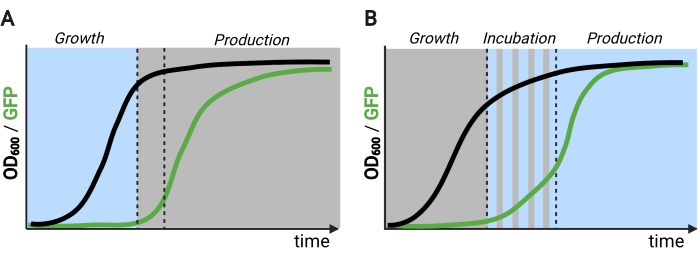
Figura 2: Fermentazioni a due e tre fasi nel tempo. (A) Le fermentazioni bifase operate con circuiti invertiti consistono in una fase di crescita guidata dalla luce e una fase di produzione scura. Nella fase di crescita, la biomassa si accumula mentre il percorso di produzione rimane represso. Al raggiungimento dell’OD600 desiderato, le cellule vengono spostate al buio per regolarsi metabolicamente prima di essere risospese in mezzi freschi per la fase di produzione. (B) In un processo in tre fasi, le fasi di crescita, incubazione e produzione sono definite da programmi di luce unici, che possono consistere in un periodo di crescita scuro, incubazione pulsata e fase di produzione completamente illuminata. Figura creata con Biorender. Fare clic qui per visualizzare una versione più grande di questa figura.
Sono stati inoltre sviluppati circuiti optogenetici per il controllo dinamico della produzione chimica e proteica in E. coli. I circuiti OptoLAC controllano il repressore batterico LacI utilizzando il circuito pDawn sensibile alla luce, basato sul sistema bicomponente YF1/FixJ6 (Figura 3). Simile a OptoINVRT5, i circuiti OptoLAC sono progettati per reprimere l’espressione genica nella luce e indurla al buio. I livelli di espressione che utilizzano circuiti OptoLAC possono eguagliare o superare quelli raggiunti con l’induzione standard isopropil β-d-1-tiogalattopoliranoside (IPTG), mantenendo così la forza dell’induzione chimica offrendo al contempo una maggiore sintonizzazione e reversibilità6. Pertanto, i circuiti OptoLAC consentono un efficace controllo optogenetico per l’ingegneria metabolica in E. coli.
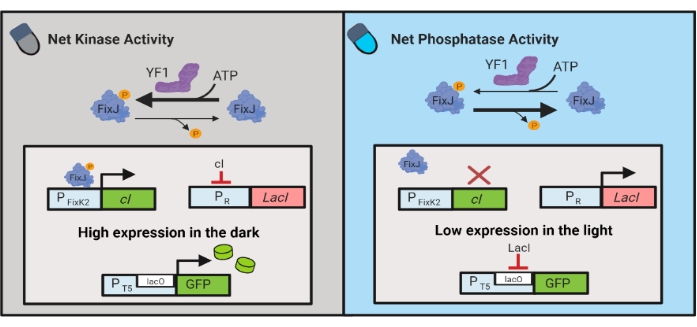
Figura 3: Circuiti OptoLAC per il controllo dinamico di E. coli. I circuiti OptoLAC adattano il sistema pDawn e l’operone lac per ottenere l’attivazione al buio e la repressione alla luce. Al buio, YF1 fosforila FixJ, che quindi attiva il promotore PFixK2 per esprimere il repressore cI . Il repressore cI impedisce l’espressione del repressore lacI dal promotore PR , che consente la trascrizione del gene di interesse da un promotore contenente lacO. Al contrario, la luce blu riduce l’attività della chinasi netta YF1, invertendo la fosforilazione di FixJ e quindi l’espressione di cI , che dereprime l’espressione di lacI e impedisce l’espressione dal promotore contenente lacO. La figura è stata modificata da Lalwani et al.6. Fare clic qui per visualizzare una versione più grande di questa figura.
Descriviamo qui i protocolli di base per le fermentazioni controllate dalla luce di S. cerevisiae ed E. coli per la produzione chimica o proteica. Sia per i lieviti che per i batteri, ci concentriamo innanzitutto sulle fermentazioni con una fase di crescita guidata dalla luce e una fase di produzione indotta dall’oscurità abilitata dai circuiti OptoINVRT e OptoLAC. Successivamente, descriviamo un protocollo per una fermentazione a tre fasi (crescita, induzione, produzione) controllata dalla luce abilitata dai circuiti OptoAMP. Inoltre, descriviamo come aumentare le fermentazioni optogeneticamente controllate dalle micropiastre ai bioreattori su scala di laboratorio. Con questo protocollo, miriamo a fornire una guida completa e facilmente riproducibile per eseguire fermentazioni controllate dalla luce per la produzione chimica o proteica.
