


Summary
ここでは、フラビンモノヌクレオチド(FMN)の光分解時に生成される活性酸素種を低強度の青色および紫色光照射下で不活化する病原菌を不活化するプロトコルを提示する。FMN光分解は、衛生プロセスのための簡単で安全な方法であることが実証されています。
Abstract
リボフラビン-5'-リン酸(またはフラビンモノヌクレオチド;FMN)は可視光に敏感です。活性酸素種(ROS)を含む様々な化合物は、可視光の照射によるFMN光分解から生成することができる。FMN光分解から発生するROSは、黄色ブドウ球菌(黄色ブドウ球菌)などの病原菌を含む微生物に有害です。本稿では、例として、可視光照射下でFMNが関与する光化学反応を介して黄色ブドウ球菌を不活性化するためのプロトコルを紹介します。FMN光分解中に生成されたスーパーオキシドラジカルアニオン()は、ニトロブルーテトラゾリウム(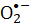 NBT)還元を介して評価されます。反応性種に起因する黄色ブドウ球菌の微生物生存率を使用して、プロセスの有効性を決定しました。細菌の不活化率はFMN濃度に比例します。紫色光は青色光照射よりも黄色ブドウ球菌の不活性化に効率的ですが、赤色または緑色の光はFMN光分解を促進しません。本稿では、衛生プロセスのための簡単で安全な方法としてのFMN光分解を示しています。
NBT)還元を介して評価されます。反応性種に起因する黄色ブドウ球菌の微生物生存率を使用して、プロセスの有効性を決定しました。細菌の不活化率はFMN濃度に比例します。紫色光は青色光照射よりも黄色ブドウ球菌の不活性化に効率的ですが、赤色または緑色の光はFMN光分解を促進しません。本稿では、衛生プロセスのための簡単で安全な方法としてのFMN光分解を示しています。
Introduction
リボフラビン-5′-リン酸(FMN)は、リビチル側鎖のリボフラビン5'位でのリン酸化によって形成され、エネルギーを生成するための多数の細胞プロセスのためにすべてのフラボタンパク質によって必要とされます。また、人体のいくつかの機能のためのビタミンの役割を果たします1。FMNは、リボフラビン2よりも約200倍水に溶けやすい。
細菌の抗菌光力学的不活性化(aPDI)は、細菌耐性の様式に依存しないため、細菌3,4に対する耐性を制御するための効率的な方法です。臨床的には、aPDIは、多耐性菌による院内皮膚の感染を減少させるために軟部組織感染症を治療するために使用されます5,6,7,8,9。aPDIはまた、活性酸素種(ROS)を生成することによって細胞死を引き起こす。スーパーオキシドラジカル()、一重項酸素、ヒドロキシルラジカル( •OH)、およびペルオキシルラジカルなどのROSは、活性酸素10,11,12を含み、通常は反応性13であるフリーラジカルまたは分子です。ROSによって引き起こされるDNA損傷と同様に、膜過酸化および内皮細胞の破壊もまた、細胞12におけるROSに起因する有害な生化学反応である。
•OH)、およびペルオキシルラジカルなどのROSは、活性酸素10,11,12を含み、通常は反応性13であるフリーラジカルまたは分子です。ROSによって引き起こされるDNA損傷と同様に、膜過酸化および内皮細胞の破壊もまた、細胞12におけるROSに起因する有害な生化学反応である。
病原性細菌に対するaPDIの使用は、塩化メチルチオニニウム14、PEI-ce6コンジュゲート15、ポルフィリン16、二酸化チタン17、トルイジンブルーO18、および酸化亜鉛ナノ粒子19などの化合物の存在下で微生物を不活性化するための可視光源またはUV光源を含む。トルイジンブルーOおよびメチレンブルーはフェノチアジニウム色素であり、メチレンブルーは毒性を有する。酸化亜鉛ナノ粒子とUV照射は、健康と環境への悪影響に関連しています。そのため、可視照射下での光分解による信頼性が高く、安全で、単純な光増感剤の開発は、さらなる研究に値します。
微量栄養素であるリボフラビンまたはFMNは毒性がなく、実際に食品の製造または給餌に使用されます20。FMNとリボフラビンはどちらも光照射に非常に敏感です2。UV1,2,21,22,23および青色光照射10,24の下で、これら2つの化合物は励起状態を達成する。光分解によって生成される活性化リボフラビンまたはFMNは、そのトリプレット状態に促進され、ROSが同時に生成されます2,25。Kumarらは、UV光によって活性化されたリボフラビンが病原性微生物におけるDNAのグアニン部分への傷害の増加を選択的に引き起こすことを報告した26。UV光の照射下で、光力学的に活性化されたリボフラビンは、二本鎖DNAにおいて、酸化ストレスのバイオマーカーである8-OH-dGの生成を促進することが実証されている27。黄色ブドウ球菌および大腸菌は、リボフラビンまたはFMN光分解中にROSによって不活性化されることが報告されています10、24、28。著者らによる以前の研究では、リボフラビンとFMNが関与する光分解反応が、クリスタルバイオレット、トリアリールメタン染料、および生成する抗菌剤を減少させ、クリスタルバイオレット28の抗菌能力のほとんどを排除することが示されました。フラビンアデニンジヌクレオチドまたはFMNに青色光を照射すると、得られたROSは、インビトロでの中毒のためにHeLa細胞にアポトーシスを生じます29。リボフラビンの存在下での光化学的処理を用いて、Cuiらはそれらの増殖およびサイトカイン産生を阻害することによってリンパ球を不活性化した30。
リボフラビンの光分解は、UV 10,24による血液病原体の不活性化に使用されるが、UV光照射下で血液成分が損なわれ得る30。また、UVに曝露された血小板は、それらの膜上の活性化マーカーP-セレクチンおよびLIMP-CD63の性能を徐々に増強することが報告されています。UVおよび高強度照射の細胞毒性を調べる必要があり、可視光を含むFMN光反応中に複雑で安全な光増感剤が非常に役立ちます。
より短い波長の光はより多くのエネルギーを持ち、細胞に深刻な損傷を引き起こす可能性がはるかに高くなります。しかしながら、適切な光増感剤の存在下では、低強度の紫色光の照射は病原性微生物を阻害することができる。したがって、紫色光を照射したときのFMNによる光増感と生成は、FMN光分解からのROSが細菌の 不活性化を増加させる経路を決定するために研究することが重要です。
抗菌剤の制御は一般的な問題であり、新しい抗生物質の開発には数十年かかることがよくあります。紫色光を照射した後、FMNによって媒介される光不活性化は、環境病原菌を全滅させることができる。この研究は、FMN光分解を促進し、したがってaPDIを生成するために紫色光を使用するin vitroで効果的な抗菌プロトコルを提示します。黄色ブドウ球菌の微生物生存率は、FMN誘発性aPDIの実現可能性を決定するために使用されます。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 光分解システムのセットアップ
- 図1に示すように、不透明なプラスチックカップ(8cm×7cm)の内側に6個の発光ダイオード(LED)(DC12V)を取り付けて、光分解システム31を確立する。
- 反応物(下記参照)をガラス試験管(直径13 mm、高さ100 mm)に加え、 図1に示すようにチューブをカップの上部に固定します。実験セットアップを25 ± 3°Cの安定した温度の部屋に置きます。
- 赤外線温度計による光分解反応全体のテストユニットの温度を監視および記録します。
注意: 図2 は、校正されたUV-Vis光学分光計を使用して記録された、青、緑、赤、および紫の光の発光スペクトルを示しています。研究で使用された青、緑、赤、紫のLEDライトの吸光度の最大値に対応する波長は、それぞれ465、529、632、および405nmです。LEDチップは、照射実験中に装置を加熱することができます。したがって、各実験の光反応システム全体は、温度が25±3°Cに一定に保たれた部屋に置かれました。
2. FMNの光分解に及ぼす光波長の影響(図3)
- 100 mM リン酸カリウムバッファー(PB)中の0.1 mM FMN溶液を調製します(pH 7.8)。FMNサンプル(各3 mL)を青、緑、赤、または紫の光に10 W / m2 の強度で5分間さらします。コントロールとして、3 mLのFMN溶液を暗所に保ちます。
- UV/可視分光光度計を使用して、照射されたFMNサンプルの吸光度を250〜750nmの範囲で記録します。
3.ニトロブルーテトラゾリウム(NBT)還元法を検出します(図4)。
注:ここで使用したNBT還元法は、リボフラビン光反応のアッセイからわずかに変更されました。NBT還元法は、FMN光分解から発生するレベルを評価するために使用される。光化学的に励起されたFMNは、まずL-メチオニンによってセミキノンに還元され、セミキノンは電子を酸素に供与して31を生じさせる。生成された状態では、NBTをホルマザンに還元し、ホルマザンは560nmに特徴的な吸光度を有する32。
- 109.3 mgのL-メチオニンを73.3 mLのPB(100 mM;pH 7.8)に加えます。溶液をボルテックスしてL-メチオニンを溶解する。
- L-メチオニンが完全に溶解したら、10 mgのNBT粉末と1.53 mLの0.117 mM FMNを溶液に加えます。適用した反応溶液(すなわち、FMN、L-メチオニン、およびNBTの混合物)1 mLごとに、FMN、メチオニン、およびNBTの最終濃度はそれぞれ2.4 x 10-6 M、1.0 x 10-2 M、および1.6 x 10-4 Mです。
- 反応溶液(1 mL)を青色または紫色のLED光照射に10 W / m2 で1〜5分間さらします。
- 光化学反応によって生成された種を(560 nmの吸光度から)検出
 し、NBTを減少させ、ホルマザンを生成します。
し、NBTを減少させ、ホルマザンを生成します。
4. FMN光分解後の 黄色ブドウ球菌 の生存率(図5)
- 黄色ブドウ球菌(BCRC 10451)のコロニーを培養プレートから15 mLスクリューキャップ試験管に採取した10 mLのLysogenyブロスに伝達します。振とう機で37°Cで16時間培養する。
- 0.5 mLの培養液を1.5 mLの遠沈管に移します。滅菌水を遠沈管に加え、培養液を600 nmで0.5の光学密度(OD600)(~6 x 107 CFU/mL)に希釈します。
- 培養液0.5 mLを1.5 mL遠沈管に移し、14,000 x g で10分間遠心分離し、上清をデカントして細胞ペレットを得た。
- ステップ4.3と同様に得られた細胞ペレットに1 mLのFMN緩衝溶液(PB溶液30、60、および120 μM FMN)を加え、ボルテックスします。照射コントロールの場合は、PBを1mL単独で添加します。
- 30 μM FMNとPBのみを含む生菌細胞液を各1 mLのガラス管に移し、紫色光を10 W/m2 で30分間照射します。
- 30、60、120 μM FMNとPBのみを含む生菌細胞液を各1 mLのガラス管に移し、20 W/m2 の青色光を120分間照射します。120 μM FMNを含む生菌細胞溶液1 mLをガラス管に別のガラス管を設置し、20 W/m2 の青色光を60分間照射します。
- 手順4.5および4.6の説明に従って試験管をセットアップし、厚いアルミホイルで覆います。これらのチューブはダークコントロールとして機能します。
- 照射チャンバー(およびダークコントロール)を9〜1°Cの冷蔵室に保管し±30〜120分の照射期間中。
注:カップ内に配置されたLEDチップは、照射実験中に光反応システムを加熱する可能性があるため、LEDライトから放出される熱を無視することはできません。したがって、実験は9 ± 1°Cに維持された冷蔵室で実施されました。 - 照射後、各反応液から0.2 mLをルリア寒天(LA)プレートに移します。L字型のガラス棒でプレート上に細菌を広げ、37°Cで一晩インキュベートします。
- 一晩の成長後の 黄色ブドウ球 菌の生存可能なプレート数と不活性化率を計算します。
注: 黄色ブドウ球菌 の不活性化率は、[1 -I / D] ×100%に等しい減少率として計算され、 I と D はそれぞれ照射されたサンプルとダークコントロールのCFUの数を示します。減少率は、不活性化率の負の値として定義されます。
5. 黄色ブドウ球菌の検出(図6)
- ステップ4で説明したように 黄色ブドウ球菌 サンプルを準備します。
- サンプルの細菌密度を滅菌水で600 nmで0.5の光学密度に希釈します(OD600、~6 x 106 CFU/mL)。0.5 mLの培養液を1.5 mLの遠沈管に移します。14,000 x g で10分間遠心分離し、上清をデカントして細胞ペレットを得た。
- 75 mL PBに0.1093 gのL-メチオニン、0.1 gのNBT、および25 mLのFMN(400、240、または120 μM)を追加します。適用される反応物1mLごとに、FMN、メチオニン、およびNBTの最終濃度はそれぞれ100(60または30)x 10-6 M、7.3 x 10-3 M、および1.2 x 10-3 Mです。
- 各反応剤溶液1mLを(すなわち、FMN濃度を変化させて)ステップ5.2のように得られた細胞ペレットに加える。紫色光を10W/m2 で10分間照射する。
- 混合物を14,000 x g で10分間遠心分離し、上清をデカントします。ペレットを1 mLのジメチルスルホキシド(DMSO)に再懸濁して、還元されたNBTを抽出します。560 nmの吸光度から産生 種を検出します。
6.統計分析
- データを3つの独立した検定の平均±標準偏差(SD)として表します。
- 等分散2標本 t検定を適用して、2つの測定値が異なるかどうかを評価します。0.05< p値は統計的に有意と見なされます。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
FMNに対する光波長の影響
青、緑、赤、紫のLEDを使用して照射された0.1 mM FMNの吸光度スペクトルを 図3に示します。ダークコントロールのFMNには2つのピーク(372nmと444nm)があります。緑と赤の光は、スペクトルの変化が重要ではないため、効果はありません。一方、444 nmのFMNの吸光度は、10 W/m2 で5分間青色光照射後、紫色光をそれぞれ約19%、34%減少させ、青色/紫色光の照射がFMN光分解を増加させることが示唆された。
の検出  青色または紫色の光を使用して照射されたFMNから
青色または紫色の光を使用して照射されたFMNから
NBT化は、32の形成を決定するために使用された。FMNは、短時間の放射線でも可視光や紫外線に非常に敏感です。 FMNの光分解中の中間光化学反応から形成されます。図4は、FMNがNBT還元に及ぼす光化学的効果は、青色または紫色の光照射に時間依存的に照射されたFMNから形成されるため、照射時間に依存することを示しています。ただし、青色光と紫色光の平均光分解効果は、図4に示すように同様です。
黄色ブドウ球菌の生存率に対するFMN光分解の影響
黄色ブドウ球菌の生存率に対する紫色または青色光照射に曝露されたFMNの影響が決定されました。青色または紫色の光照射を用いたFMN光分解は、黄色ブドウ球菌の有意な不活性化をもたらす。FMNの濃度が高いほど、図5Aに示すように、20W/m2で120分間青色光を照射した黄色ブドウ球菌の不活化率が高くなります。黄色ブドウ球菌では、20 W/m2の青色光照射下で120分間、それぞれ0、30、60、および120 μM FMNを使用して、26%、39%、43%、および97%の不活化率が達成されました。図5Aに示すように、60 μM FMNを20 W/m 2で120分間照射した場合(エネルギー線量、14.4 J/cm 2)、または120 μM FMNを20 W/m 2で60分間照射した場合(エネルギー線量、 7.2 J/cm2)。一方、図5Bに示すように、黄色ブドウ球菌では、10 W/m 2で30分間(エネルギー線量1.8 J/cm2)の紫色光照射を行い、30 μM FMNなしと使用した場合で、それぞれ53%と96%の不活化率が達成されました。したがって、青色または紫色の光照射にさらされたFMNは、細菌の不活性化を増加させることにより、黄色ブドウ球菌の生存率に大きな影響を与えます。紫色光照射で処理されたFMNは、黄色ブドウ球菌の不活性化においてより効率的である。
の検出 紫色光照射下でのFMN処理黄色ブドウ球菌
紫色光照射下でのFMN処理黄色ブドウ球菌の産生をNBT低減法を用いて測定した。560nmにおける吸光度値を用いて、産生の程度を評価した。黄色ブドウ球菌の生成に対するFMNの光化学的効果は、FMN濃度に比例します(図6)。FMN処理なしの黄色ブドウ球菌の560 nmの吸光度値は、10 W/m2の紫色光照射下で10分間および暗所でそれぞれ0.31および0.07です(図6)。このことは、紫色光照射後の未処理の黄色ブドウ球菌ではほとんど産生されなかったことを示唆している。100 μM FMNで処理した黄色ブドウ球菌のそれぞれの560 nmの吸光度値は、10 W / m2の紫色光照射下で10分間および暗所でそれぞれ1.36および0.08です。したがって、FMN処理黄色ブドウ球菌のNBT還元に対する光化学的効果は、豊富に生成されるように、紫色照射下で増加します。

図1:光反応系の模式図。 この図の拡大版を表示するには、ここをクリックしてください。

図2:この研究における青、緑、赤、紫のLEDライトの発光スペクトル。 青、緑、赤、紫の光の発光極大はそれぞれ465、529、632、405nmであり、ハーフハイト(W1/2)でのスペクトル幅はそれぞれ26、31、14、12 nmです。 この図の拡大版を表示するには、ここをクリックしてください。

図3:さまざまな波長のLEDで10 W/m2 で5分間処理したFMNの吸光度スペクトル。 この図の拡大版を表示するには、ここをクリックしてください。

図4:FMN光分解に対する10 W / m2 での青色および紫色光照射の影響を決定するためのNBT還元。 この図の拡大版を表示するには、ここをクリックしてください。

図5:黄色ブドウ球菌の生存率に対するFMN光分解の影響。減少率は、(A)FMNを20W/m2で120分間照射し、(B)紫色光を10W/m2で30分間照射したFMNで処理した黄色ブドウ球菌の不活化率の負の値と定義し、FMN濃度と照射時間を図の上部に示します。なお、黄色ブドウ球菌では、60 μM FMNを20 W/m2の青色光照射で120分間、または120 μM FMNを20 W/m2の青色光照射で60分間使用した場合(A)に有意差はありません。結果は、n = 3の場合、SD±平均値で表されます。*p < 0.05;**p < 0.01;p < 0.0001この図の拡大版を表示するには、ここをクリックしてください。

図6:10 W/m2で10分間紫色光を照射した黄色ブドウ球菌の発生に及ぼす FMN濃度の影響。全体として、FMNの非存在下で黄色ブドウ球菌ではほとんど生成されませんが、紫色光の照射は、暗い対照(PB(暗い))と比較して照射されたサンプル(PB(紫色光))でわずかに増加します。結果は、n = 3の場合、SD±平均値で表されます。* p < 0.05;** p < 0.01; p < 0.0001この図の拡大版を表示するには、ここをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
光増感剤は、化合物の光化学反応を高めてROSを生成します。病原性微生物は、光増感剤の存在下で光照射することによって不活性化することができる。本研究では、外因性光増感剤FMNの紫色光照射により発生するROSに起因する 黄色ブドウ球菌 のaPDIを決定した。
図3に示すように、FMNの場合、紫色または青色光を用いた5分間の照射後に444nmの吸光度は有意に低下し、緑色または赤色光の照射下でのFMN吸光度に大きな変化はありません。NBT化は、電荷移動プロセス10、33を介したの形成を決定するために使用される。著者らによる以前の研究では、青色、緑色、または赤色光照射下でFMNを使用して、NBT還元を使用した形成を決定しました。青色光の照射は莫大な量を生成するのに対し、緑色光および赤色光の光分解効率は青色光によって達成されるものの3%未満である10。これらの結果は、FMNに青色または紫色の光照射を照射すると電荷移動プロセスが生じ、緑色または赤色の光の照射によって大きな変化がないことを示しています。
著者らによる以前の研究34 は、FMN光分解のメカニズムが一次FMN光分解反応(式1)で説明できることを示しました。
 (1),
(1),
ここで、1FMN* と 3FMN* はそれぞれ励起 FMN の一重項状態と三重項状態です。これらの光誘起FMNの光分解反応機構は、おそらくトリプレット-トリプレット消滅またはトリプレット基底状態消光プロセスによって開始される35,36,37,38。FMN の基底状態 (FMN0) は、式 2 に示すように、3FMN* に電子を供給し、半還元型 (FMN•-) および半酸化型 (FMN•+) を生成する電子供与体です。
 (2)
(2)
FMNの光分解経路は、光増感に起因します。FMNは光分解反応によって電子励起状態FMNに促進され、FMN•が電子を失うと中性FMNが還元されます(以下の式3-8を参照)。
 (3)
(3)
 (4)
(4)
 (5),
(5),
ここで、FMN•+ はラジカルカチオン種です。
FMN•-とO2(3Σg-)との相互作用により、反応性スーパーオキシドラジカルが生成され、続いてヒドロペルオキシルラジカルHOO•が生成され、プロトンとの反応で生成され、最終的にFMN0の分解につながります。
 (6)
(6)
 (7)
(7)
 (8),
(8),
ここで、FMN•- はラジカルアニオンです。
本研究では、FMNの非存在下での黄色ブドウ球菌の不活化率は、紫色光照射10W/m2で30分間で53%、青色光照射20W/m2で120分間照射すると26%となり、紫色光照射よりも失活する細菌が少ないことが示唆されました(図5)。黄色ブドウ球菌のポルフィリンは405±5 nmで最大吸光度を持ち、照射後に細菌の不活性化を引き起こすが、外因性の光増感剤なしで430nmより長い波長で照射しても黄色ブドウ球菌細胞を不活性化しないことが以前に報告されている39。
本著者らによる以前の研究では、大腸菌および黄色ブドウ球菌が、リボフラビンまたはFMN 10,24,40の光分解を介した形成によって誘導されるDNA切断によって不活性化されることが示された。黄色ブドウ球菌の不活化率は、120 μM FMNを20 W/m 2の青色光照射(エネルギー線量14.4 J/cm 2)で120分間、30 μM FMNを10 W/m 2の紫色光照射(エネルギー線量1.8 J/cm 2)で30分間使用した場合、それぞれ97%と96%です(図5)。したがって、紫色光の低エネルギー線量は、青色光と比較してより低いFMN濃度およびより短い照明期間の存在下で黄色ブドウ球菌を不活性化することができるので、紫色光照射は黄色ブドウ球菌を不活性化するのにより効果的である。紫色光によって達成されるFMN光分解の程度は青色光と比較して高いが(図3)、青色光照射および紫色光照射下でのNBT還元に対するFMNの平均光分解効果は類似している(図4)。
大腸菌は一般的なグラム陰性菌です。青色光照射によるリボフラビンまたはFMN光分解は、FMN 10,24を使用した場合、ROS10,24,41を介して大腸菌を不活性化し、不活化率は96%です。一方、黄色ブドウ球菌はグラム陽性菌であり、ここで示されているように、紫色光照射下でのFMN光分解によって効果的に不活性化されます(図5)。したがって、青色または紫色の光照射下でのFMN光分解反応は、ROSを生成し、グラム陰性菌とグラム陽性菌の両方を不活性化する可能性があります。
しかし、FMNの非存在下では、紫色光照射に曝露された黄色ブドウ球菌ではほとんど産生されないため(図6)、したがって、内因性光増感剤は、FMNなどの外因性光増感剤の存在下での効果と比較して、紫色光照射下での黄色ブドウ球菌の生存率をわずかに低下させるだけです。同じ条件下で、紫色光照射にさらされたFMNは、黄色ブドウ球菌の生存率を低下させ、96%の不活性化率をもたらし、これは内因性細胞内光増感剤単独の場合よりもはるかに高い。紫色光照射下でのFMN処理黄色ブドウ球菌の産生は、図6に示すように有意に増加した。これらの結果は、紫色光照射にさらされたFMNが細菌不活性化のレベルの上昇を引き起こすことを示しています。
FMNは、UV1、2、21、22、23、青色10、24、および紫色光31を使用して照射すると、光励起状態を達成できます。しかしながら、紫外線および高強度短波長照射によって引き起こされる細胞毒性は考慮されなければならない42。一方、より長い波長の赤色および緑色の放射線は、FMN光分解中に有意な量のROSを生成しません10。したがって、黄色ブドウ球菌のROS誘起aPDIは、FMNに中間波長の紫色光を照射する必要がある。紫色光の半値高さ(W1/2)でのスペクトル幅は、UV吸収との重なりを避け、UV放射によるリスクを抑制するために狭くなければならない31。微量栄養素であるFMNは無害であり、人体に不可欠なビタミンです。FMNなどの適切な光増感剤を使用すると、低エネルギー紫色光照射は病原菌を抑制し、衛生プロセスの安全で効果的な手段を提供します。黄色ブドウ球菌に対するFMNの光分解反応に加えて、微生物に対するFMNの光毒性など、他の問題はさらなる調査に値するものであり、qPCRとともにRNA-seq技術を使用して研究することができます。
紫色光照射 による FMN光分解は、ROSを生成する光化学的酸化をもたらし、病原菌の不活性化を促進します。紫色光の使用は、FMN光分解における重要なステップと考えられています。現在の技術の限界は、紫外線照射によるリスクを回避するために、紫色光の波長間隔を非常に狭くしなければならないことである。現在の方法は、光ファイバーを挿入して紫色光を感染部位に向けることにより、皮下組織や筋肉の深部とともに表面の皮膚の感染治療など、将来の医療応用の可能性を秘めています。したがって、FMNを使用した光化学処理は、効果的な衛生慣行を確保するための安全で簡単な方法です。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者は利益相反を宣言しません。
Acknowledgments
著者らは、Tak-Wah Wong博士とZong-Jhe Hsieh氏の実験支援に感謝している。
Materials
| Name | Company | Catalog Number | Comments |
| Blue, green and red LED lights | Vita LED Technologies Co., Tainan, Taiwan | DC 12 V 5050 | |
| Dimethyl Sulfoxide | Sigma-Aldrich, St. Louis, MO | 190186 | |
| Infrared thermometer | Raytek Co. Santa Cruz, CA | MT4 | |
| LB broth | Difco Co., NJ | ||
| L-Methionine | Sigma-Aldrich, St. Louis, MO | 1.05707 | |
| NBT | Bio Basic, Inc. Markham, Ontario, Canada | ||
| Power supply | China tech Co., New Taipei City, Taiwan | YP30-3-2 | |
| Riboflavin 5′-phosphate | Sigma-Aldrich, St. Louis, MO | R7774 | |
| RNase | New England BioLabs, Inc. Ipswich, MA | ||
| Solar power meter | Tenmars Electronics Co., Taipei, Taiwan | TM-207 | |
| Staphylococcus aureus subsp. aureus | Bioresource Collection and Research Center (BCRC), Hsinchu, Taiwan | 10451 | |
| UV-Vis optical spectrometer | Ocean Optics, Dunedin, FL | USB4000 | |
| UV-Vis spectrophotometer | Hitachi High-Tech Science Corporation,Tokyo, Japan | U-2900 | |
| Violet LED | Long-hui Electronic Co., LTD, Dongguan, China |
References
- Jian, H. L., Cheng, C. W., Chen, L. Y., Liang, J. Y.
- Lin, Y., Eitenmiller, R. R., Landen, W. O. Riboflavin. Vitamin analysis for the health and food sciences. , CRC Press. 329-360 (2008).
- Xie, L. J., Wang, R. L., Wang, D., Liu, L., Cheng, L. Visible-light-mediated oxidative demethylation of N(6)-methyl adenines. Chemical Communications. 53 (77), 10734-10737 (2017).
- Tim, M. Strategies to optimize photosensitizers for photodynamic inactivation of bacteria. Journal of Photochemistry and Photobiology B: Biology. 150, 2-10 (2015).
- Maisch, T., et al. Photodynamic inactivation of multi-resistant bacteria (PIB) - a new approach to treat superficial infections in the 21st century. Journal of the German Society of Dermatology. 9 (5), 360-366 (2011).
- Hamblin, M. R. Antimicrobial photodynamic inactivation: a bright new technique to kill resistant microbes. Current Opinion in Microbiology. 33, 67-73 (2016).
- Del Rosso, J. Q. Oral Doxycycline in the management of acne vulgaris: Current perspectives on clinical use and recent findings with a new double-scored small tablet formulation. The Journal of Clinical and Aesthetic Dermatology. 8 (5), 19-26 (2015).
- Wong, T. W., et al. Photodynamic inactivation of methicillin-resistant Staphylococcus aureus by indocyanine green and near infrared light. Dematologica Sinica. 36 (1), 8-15 (2018).
- Yuann, J. M. P., et al. Effects of free radicals from doxycycline hyclate and minocycline hydrochloride under blue light irradiation on the deactivation of Staphylococcus aureus, including a methicillin-resistant strain. Journal of Photochemistry and Photobiology B: Biology. 226, 112370 (2022).
- Liang, J. Y., Cheng, C. W., Yu, C. H., Chen, L. Y. Investigations of blue light-induced reactive oxygen species from flavin mononucleotide on inactivation of E. coli. Journal of Photochemistry and Photobiology B: Biology. 143, 82-88 (2015).
- Yang, M. J., et al. Effects of blue-light-induced free radical formation from catechin hydrate on the inactivation of Acinetobacter baumannii, Including a carbapenem-resistant strain. Molecules. 23 (7), 1631 (2018).
- Yuann, J. M. P., et al. A study of catechin photostability using photolytic processing. Processes. 9 (2), 293 (2021).
- Yuann, J. M. P., Wang, J. S., Jian, H. L., Lin, C. C., Liang, J. Y. Effects of Clinacanthus nutans (Burm. f) Lindau leaf extracts on protection of plasmid DNA from riboflavin photoreaction. MC-Transaction on Biotechnology. 4 (1), 45-59 (2012).
- Rineh, A., et al. Attaching NorA efflux pump inhibitors to methylene blue enhances antimicrobial photodynamic inactivation of Escherichia coli and Acinetobacter baumannii in vitro and in vivo. Bioorganic and Medicinal Chemistry Letters. 28 (16), 2736-2740 (2018).
- Dai, T., et al. Photodynamic therapy for Acinetobacter baumannii burn infections in mice. Antimicrobial Agents and Chemotherapy. 53 (9), 3929-3934 (2009).
- Nitzan, Y., Ashkenazi, H. Photoinactivation of Acinetobacter baumannii and Escherichia coli B by a cationic hydrophilic porphyrin at various light wavelengths. Current Microbiology. 42 (6), 408-414 (2001).
- Tseng, C. C., et al. Altered susceptibility to the bactericidal effect of photocatalytic oxidation by TiO2 is related to colistin resistance development in Acinetobacter baumannii. Applied Microbiology and Biotechnology. 100 (19), 8549-8561 (2016).
- Boluki, E., et al. Antimicrobial activity of photodynamic therapy in combination with colistin against a pan-drug resistant Acinetobacter baumannii isolated from burn patient. Photodiagnosis and Photodynamic Therapy. 18, 1-5 (2017).
- Yang, M. Y., et al. Blue light irradiation triggers the antimicrobial potential of ZnO nanoparticles on drug-resistant Acinetobacter baumannii. Journal of Photochemistry and Photobiology B, Biology. 180, 235-242 (2018).
- Code of Federal Regulations. , Available from: https://www.gpo.gov/fdsys/pkg/CFR-2009-title21-vol6/pdf/CFR-2009-title21-vol6-sec582-5695.pdf (2022).
- Lu, C. Y., et al. Generation and photosensitization properties of the oxidized radical of riboflavin: a laser flash photolysis study. Journal of Photochemistry and Photobiology B, Biology. 52 (1-3), 111-116 (1999).
- Sato, K., et al. The primary cytotoxicity in ultraviolet-a-irradiated riboflavin solution is derived from hydrogen peroxide. The Journal of Investigative Dermatology. 105 (4), 608-612 (1995).
- Tripathi, A. K., et al. Attenuated neuroprotective effect of riboflavin under UV-B irradiation via miR-203/c-Jun signaling pathway in vivo and in vitro. Journal of Biomedical Science. 21 (1), 39 (2014).
- Liang, J. Y., et al. Blue light induced free radicals from riboflavin on E. coli DNA damage. Journal of Photochemistry and Photobiology B, Biology. 119, 60-64 (2013).
- Ottaway, P. B. The Technology of Vitamins in Food. , Chapman and Hall. London. Ch. 5 233-244 (1993).
- Kumar, V., et al. Riboflavin and UV-light based pathogen reduction: extent and consequence of DNA damage at the molecular level. Photochemistry and Photobiology. 80 (1), 15-21 (2004).
- Ito, K., Inoue, S., Yamamoto, K., Kawanishi, S. 8-Hydroxydeoxyguanosine formation at the 5'site of 5'-GG-3'sequences in double-stranded DNA by UV radiation with riboflavin. Journal of Biological Chemistry. 268 (18), 13221-13227 (1993).
- Liang, J. Y., Yuann, J. M. P., Hsie, Z. J., Huang, S. T., Chen, C. C. Blue light induced free radicals from riboflavin in degradation of crystal violet by microbial viability evaluation. Journal of Photochemistry and Photobiology B, Biology. 174, 355-363 (2017).
- Yang, M. Y., Chang, C. J., Chen, L. Y. Blue light induced reactive oxygen species from flavin mononucleotide and flavin adenine dinucleotide on lethality of HeLa cells. Journal of Photochemistry and Photobiology B, Biology. 173, 325-332 (2017).
- Cui, Z., Huang, Y., Mo, Q., Wang, X., Qian, K. Inactivation of lymphocytes in blood products using riboflavin photochemical treatment with visible light. Photochemistry and Photobiology. 84 (5), 1195-1200 (2008).
- Wong, T. W., Cheng, C. W., Hsieh, Z. J., Liang, J. Y. Effects of blue or violet light on the inactivation of Staphylococcus aureus by riboflavin-5′-phosphate photolysis. Journal of Photochemistry and Photobiology B: Biology. 173, 672-680 (2017).
- Russell, L. F., Vanderslice, J. T. Comprehensive review of vitamin B2 analytical methodology. Journal of Micronutrient Analysis. 8, 257-310 (1990).
- Cheng, C. W., Chen, L. Y., Chou, C. W., Liang, J. Y. Investigations of riboflavin photolysis via coloured light in the nitro blue tetrazolium assay for superoxide dismutase activity. Journal of Photochemistry and Photobiology B: Biology. 148, 262-267 (2015).
- Huang, S. T., et al. Effects of 462 nm light-emitting diode on the inactivation of Escherichia coli and a multidrug-resistant by tetracycline photoreaction. Journal of Clinical Medicine. 7 (9), 278 (2018).
- Barua, M. G., Escalada, J. P., Bregliani, M., Pajares, A., Criado, S. Antioxidant capacity of (+)-catechin visible-light photoirradiated in the presence of vitamin B2. Redox Report: Communications in Free Radical Research. 22 (6), 282-289 (2017).
- Castillo, C., Criado, S., Díaz, M., García, N. A. Riboflavin as a sensitiser in the photodegradation of tetracyclines. Kinetics, mechanism and microbiological implications. Dyes and Pigments. 72 (2), 178-184 (2007).
- Huvaere, K., Sinnaeve, B., Van Bocxlaer, J., Skibsted, L. H. Flavonoid deactivation of excited state flavins: reaction monitoring by mass spectrometry. Journal of Agricultural and Food Chemistry. 60 (36), 9261-9272 (2012).
- Massad, W. A., Bertolotti, S., Garcia, N. A. Kinetics and mechanism of the vitamin B2-sensitized photooxidation of isoproterenol. Photochemistry and Photobiology. 79 (5), 428-433 (2004).
- Maclean, M., MacGregor, S. J., Anderson, J. G., Woolsey, G. High-intensity narrow-spectrum light inactivation and wavelength sensitivity of Staphylococcus aureus. FEMS Microbiology Letters. 285 (2), 227-232 (2008).
- Huang, S. T., et al. The influence of the degradation of tetracycline by free radicals from riboflavin-5'-phosphate photolysis on microbial viability. Microorganisms. 7 (11), 500 (2019).
- Maisch, T., et al. Fast and effective photodynamic inactivation of multiresistant bacteria by cationic riboflavin derivatives. PLoS One. 9 (12), 111792 (2014).
- Grijzenhout, M., et al. Ultraviolet-B irradiation of platelets induces a dose-dependent increase in the expression of platelet activation markers with storage. British Journal of Haematology. 83 (4), 627-632 (1993).
