


Summary
Här presenterar vi ett protokoll för att inaktivera patogena bakterier med reaktiva syrearter som produceras under fotolys av flavinmononukleotid (FMN) under bestrålning av blått och violett ljus med låg intensitet. FMN-fotolys har visat sig vara en enkel och säker metod för sanitära processer.
Abstract
Riboflavin-5'-fosfat (eller flavinmononukleotid; FMN) är känslig för synligt ljus. Olika föreningar, inklusive reaktiva syrearter (ROS), kan genereras från FMN-fotolys vid bestrålning med synligt ljus. ROS som genereras från FMN-fotolys är skadliga för mikroorganismer, inklusive patogena bakterier som Staphylococcus aureus (S. aureus). Denna artikel presenterar ett protokoll för att inaktivera S. aureus, som ett exempel, via fotokemiska reaktioner som involverar FMN under bestrålning av synligt ljus. Superoxidradikalanjonen () som genereras under FMN-fotolysen utvärderas via nitroblå tetrazoliumreduktion (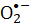 NBT). Den mikrobiella livskraften hos S. aureus som tillskrivs reaktiva arter användes för att bestämma processens effektivitet. Den bakteriella inaktiveringshastigheten är proportionell mot FMN-koncentrationen. Violett ljus är effektivare för att inaktivera S. aureus än bestrålning av blått ljus, medan det röda eller gröna ljuset inte driver FMN-fotolys. Denna artikel visar FMN-fotolys som en enkel och säker metod för sanitära processer.
NBT). Den mikrobiella livskraften hos S. aureus som tillskrivs reaktiva arter användes för att bestämma processens effektivitet. Den bakteriella inaktiveringshastigheten är proportionell mot FMN-koncentrationen. Violett ljus är effektivare för att inaktivera S. aureus än bestrålning av blått ljus, medan det röda eller gröna ljuset inte driver FMN-fotolys. Denna artikel visar FMN-fotolys som en enkel och säker metod för sanitära processer.
Introduction
Riboflavin-5′-fosfat (FMN) bildas genom fosforylering vid riboflavin 5′-positionen i ribityl-sidokedjan och krävs av alla flavoproteiner för många cellulära processer för att generera energi. Det spelar också rollen som vitamin för vissa funktioner i människokroppen1. FMN är cirka 200 gånger mer lösligt i vatten än riboflavin2.
Den antibakteriella fotodynamiska inaktiveringen (aPDI) av bakterier är ett effektivt sätt att kontrollera resistens mot bakterier 3,4 eftersom det inte beror på läget för bakteriell resistens. Kliniskt används aPDI för att behandla mjukdelsinfektioner för att minska infektion i nosokomial hud på grund av multiresistenta bakterier 5,6,7,8,9. aPDI producerar också celldöd genom att generera reaktiva syrearter (ROS). ROS, såsom superoxidradikaler (), singlet syre, hydroxylradikaler ( •OH) och peroxylradikaler, är fria radikaler eller molekyler som innehåller reaktivt syre 10,11,12 och är normalt reaktiva 13. I likhet med DNA-skador som orsakas av ROS är membranperoxidation och förstörelse av endotelceller också negativa biokemiska reaktioner som tillskrivs ROS i celler12.
•OH) och peroxylradikaler, är fria radikaler eller molekyler som innehåller reaktivt syre 10,11,12 och är normalt reaktiva 13. I likhet med DNA-skador som orsakas av ROS är membranperoxidation och förstörelse av endotelceller också negativa biokemiska reaktioner som tillskrivs ROS i celler12.
Användningen av aPDI för patogena bakterier involverar en synlig eller UV-ljuskälla för att inaktivera mikroorganismer i närvaro av kemiska föreningar, såsom metyltioninklorid 14, PEI-c e6-konjugat 15, porfyrin16, titandioxid 17, toluidinblå O 18 och zinkoxidnanopartiklar 19. Toluidinblå O och metylenblå är fenotiaziniumfärger och metylenblått har toxiska egenskaper. Nanopartiklar av zinkoxid och UV-bestrålning är kopplade till negativa hälso- och miljöeffekter. Som sådan förtjänar utnyttjandet av en pålitlig, säker och enkel fotosensibiliserare via fotolys under synlig bestrålning ytterligare studier.
Mikronäringsämnet, riboflavin eller FMN, är inte giftigt och används verkligen för livsmedelstillverkning eller utfodring20. Både FMN och riboflavin är mycket känsliga för ljusbestrålning2. Under UV 1,2,21,22,23 och bestrålning av blått ljus 10,24 uppnår dessa två föreningar ett exciterat tillstånd. Det aktiverade riboflavinet eller FMN som produceras genom fotolys befordras till sitt tripletttillstånd och ROS genereras samtidigt 2,25. rapporterade att riboflavin aktiverat av UV-ljus selektivt orsakar ökad skada på guanindelen av DNA i patogena mikroorganismer26. Under bestrålning med UV-ljus har fotodynamiskt aktiverat riboflavin visat sig främja genereringen av 8-OH-dG, som är en biomarkör för oxidativ stress, i dubbelsträngat DNA27. Det rapporteras att S. aureus och E. coli deaktiveras av ROS under riboflavin eller FMN fotolys 10,24,28. En tidigare studie av författarna visade att de fotolytiska reaktionerna som involverar riboflavin och FMN minskar kristallviolett, ett triarylmetanfärgämne och ett antibakteriellt medel som genererar , och eliminerar det mesta av den antimikrobiella förmågan hos kristallviolett28. När flavinadenindinukleotid eller FMN bestrålas av blått ljus producerar den resulterande ROS apoptos i HeLa-celler för deras förgiftning in vitro29. Med hjälp av fotokemisk behandling i närvaro av riboflavin, cui et al. inaktiverade lymfocyter genom att hämma deras proliferation och cytokinproduktion30.
Fotolysen av riboflavin används för inaktivering av blodpatogen med UV 10,24, men blodkomponenter kan försämras vid bestrålning av UV-ljus30. Det rapporteras också att blodplättar som utsätts för UV gradvis förbättrar prestandan hos aktiveringsmarkörerna P-selectin och LIMP-CD63 på deras membran. Cytotoxiciteten hos UV och högintensiv bestrålning måste undersökas och en fotosensibiliserare som är okomplicerad och säker under en FMN-fotoreaktion med synligt ljus skulle vara till stor nytta.
Ljus med kortare våglängder har mer energi och är mycket mer benägna att orsaka allvarliga skador på celler. I närvaro av en lämplig fotosensibiliserare kan emellertid bestrålning med lågintensivt violett ljus hämma patogena mikroorganismer. Fotosensibiliseringen och genereringen av av FMN när den bestrålas med violett ljus är därför viktig att studera för att bestämma vägen genom vilken ROS från FMN-fotolys ökar inaktiveringen av bakterier.
Antimikrobiell kontroll är en vanlig fråga och utvecklingen av nya antibiotika tar ofta årtionden. Efter bestrålning med violett ljus kan fotoinaktivering som förmedlas av FMN förinta miljöpatogena bakterier. Denna studie presenterar ett effektivt antimikrobiellt protokoll in vitro med violett ljus för att driva FMN-fotolys och därmed generera för aPDI. Den mikrobiella livskraften hos S. aureus används för att bestämma genomförbarheten av FMN-inducerad aPDI.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Fotolyssystem inställning
- Montera sex lysdioder (LED) (DC 12 V) på insidan av en ogenomskinlig plastkopp (8 cm x 7 cm) enligt figur 1 för att upprätta ett fotolyssystem31.
- Tillsätt reaktanter (se nedan) i glasprovrören (13 mm i diameter och 100 mm i höjd) och säkra rören högst upp på koppen enligt figur 1. Placera experimentuppställningen i ett rum med en jämn temperatur på 25 ± 3 °C.
- Övervaka och registrera temperaturen på testenheter under de fotolytiska reaktionerna med en infraröd termometer.
OBS: Figur 2 visar emissionsspektra för blått, grönt, rött och violett ljus, registrerat med en kalibrerad optisk UV-Vis-spektrometer. Våglängderna som motsvarar absorbansmaximum för blå, grön, röd, och violetta LED-lampor som används i studien är 465, 529, 632, respektive 405 nm. LED-chipsen kan värma apparaten under bestrålningsexperiment. Hela fotoreaktionssystemet i varje experiment placerades därför i ett rum där temperaturen hölls konstant vid 25 ± 3 °C.
2. Effekten av ljusvåglängd på fotolysen av FMN (figur 3)
- Bered en 0,1 mM FMN-lösning i 100 mM kaliumfosfatbuffert (PB) (pH 7,8). Exponera FMN-prover (3 ml vardera) för blått, grönt, rött eller violett ljus med en intensitet på 10 W/m2 i 5 minuter. Förvara 3 ml FMN-lösning i mörker som en kontroll.
- Registrera absorbansen hos bestrålade FMN-prover i intervallet 250–750 nm med hjälp av en UV/synlig spektrofotometer.
3. Nitroblå tetrazolium (NBT) reduktionsmetod för detektion (figur 4).
OBS: NBT-reduktionsmetoden som används här modifierades något från analysen för riboflavinfotoreaktion. NBT-reduktionsmetoden används för att utvärdera nivån av genererad från FMN-fotolys. Det fotokemiskt exciterade FMN reduceras först av L-metionin till en semikinon, som sedan donerar en elektron till syre som ger upphov till 31. Den genererade reducerar NBT till formazan, som har en karakteristisk absorbans vid 560 nm32.
- Tillsätt 109,3 mg L-metionin till 73,3 ml PB (100 mM; pH 7,8). Vortex lösningen för att lösa upp L-metionin.
- Efter att L-metionin är fullständigt upplöst, tillsätt 10 mg NBT-pulver och 1,53 ml 0,117 mM FMN till lösningen. För varje 1 ml av reaktionslösningen (dvs. en blandning av FMN, L-metionin och NBT) som appliceras är de slutliga koncentrationerna av FMN, metionin och NBT 2,4 x 10-6 M, 1,0 x 10-2 M respektive 1,6 x 10-4 M.
- Utsätt reaktionslösningen (1 ml) för bestrålning av blått eller violett LED-ljus vid 10 W/m2 i 1-5 min.
- Upptäck
 arten (från absorbansen vid 560 nm) som produceras av den fotokemiska reaktionen, vilket minskar NBT och producerar formazan.
arten (från absorbansen vid 560 nm) som produceras av den fotokemiska reaktionen, vilket minskar NBT och producerar formazan.
4. S. aureus viabilitet efter FMN-fotolys (figur 5)
- Överför en koloni av S. aureus (BCRC 10451) från en odlad platta till 10 ml lysogenibuljong som tas i ett 15 ml skruvtäckt provrör. Odla i en shaker vid 37 °C i 16 timmar.
- Överför 0,5 ml av kulturen till ett 1,5 ml centrifugrör. Tillsätt steriliserat vatten i centrifugröret för att späda ut kulturen till en optisk densitet på 0,5 vid 600 nm (OD600) (~6 x 107 CFU/ml).
- Överför 0,5 ml av kulturen till ett 1,5 ml centrifugrör, centrifugera vid 14 000 x g i 10 minuter och dekantera supernatanten för att erhålla en cellpellet.
- Tillsätt 1 ml FMN-buffrad lösning (30, 60 och 120 μM FMN i PB) till de cellpellets som erhålls som i steg 4.3 och virvel. För bestrålad kontroll, tillsätt 1 ml PB ensam.
- Överför 1 ml vardera av livskraftiga bakteriecelllösningar som innehåller 30 μM FMN och PB ensamma till glasrör och bestråla dem med violett ljus vid 10 W / m2 i 30 min.
- Överför 1 ml vardera av livskraftiga bakteriecelllösningar innehållande 30, 60 och 120 μM FMN och PB ensamma till glasrör och bestråla dem med blått ljus vid 20 W / m2 i 120 min. Sätt upp ett annat glasrör med 1 ml livskraftig bakteriecelllösning som innehåller 120 μM FMN och bestråla det med blått ljus vid 20 W/m2 i 60 min.
- Ställ in provrör enligt beskrivningen i steg 4.5 och 4.6 och täck dem med tjocka aluminiumfolier. Dessa rör fungerar som mörka kontroller.
- Förvara bestrålningskammaren (liksom de mörka kontrollerna) i ett kylrum vid 9 ± 1 °C under bestrålningsperioden 30-120 minuter.
OBS: Värme som frigörs av LED-lampor kan inte ignoreras eftersom LED-chipsen som placeras inuti koppen kan värma fotoreaktionssystemet under bestrålningsexperiment. Experimenten utfördes därför i ett kylrum som hölls vid 9 ± 1 °C. - Efter bestrålning, överför 0,2 ml från var och en av reaktionslösningarna till en Luria agar (LA) platta. Sprid bakterierna över plattan med en L-formad glasstav och inkubera över natten vid 37 °C.
- Beräkna det livskraftiga antalet plattor och inaktiveringshastigheterna för S. aureus efter tillväxt över natten.
OBS: Inaktiveringsgraden för S. aureus beräknas som den procentuella reduktionen, som är lika med [1 -I / D] ×100%, där I och D betecknar respektive antalet CFUs i det bestrålade provet och mörk kontroll. Den procentuella minskningen definieras som ett negativt värde för inaktiveringsgraden.
5. Detektion av i S. aureus (figur 6)
- Förbered S. aureus-proverna enligt beskrivningen i steg 4.
- Späd provernas bakterietäthet med steriliserat vatten till en optisk densitet på 0,5 vid 600 nm (OD600, ~6 x 106 CFU/ml). Överför 0,5 ml av kulturen till ett 1,5 ml centrifugrör. Centrifugera vid 14 000 x g i 10 minuter och dekantera supernatanten för att få en cellpellet.
- Tillsätt 0,1093 g L-metionin, 0,1 g NBT och 25 ml FMN (400, 240 eller 120 μM) till 75 ml PB. För varje 1 ml av den applicerade reaktanten är de slutliga koncentrationerna av FMN, metionin och NBT 100 (60 eller 30) x 10-6 M, 7,3 x 10-3 M respektive 1,2 x 10-3 M.
- Tillsätt 1 ml av varje reaktantlösning (dvs. med varierande FMN-koncentrationer) till de cellpellets som erhålls enligt steg 5.2. Bestråla lösningarna med violett ljus vid 10 W/m2 i 10 min.
- Centrifugera blandningen vid 14 000 x g i 10 min och dekantera supernatanten. Återsuspendera pelleten i 1 ml dimetylsulfoxid (DMSO) för att extrahera den reducerade NBT. Upptäck de producerade arterna från absorbansen vid 560 nm.
6. Statistisk analys
- Expressdata som medelvärde ± standardavvikelse (SD) för tre oberoende tester.
- Applicera ett homoscedastiskt t-test med två prover för att utvärdera om två mätningar är olika. p-värden < 0,05 anses vara statistiskt signifikanta.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Effekt av ljusvåglängd på FMN
Absorbansspektra på 0,1 mM FMN som bestrålas med blå, gröna, röda och violetta lysdioder visas i figur 3. Det finns två toppar för FMN (372 nm och 444 nm) för den mörka kontrollen. Grönt och rött ljus har ingen effekt eftersom förändringar i spektra är obetydliga. Å andra sidan reduceras respektive absorbans av FMN vid 444 nm med cirka 19% respektive 34% efter bestrålning av blått och violett ljus vid 10 W / m2 i 5 min, vilket tyder på att bestrålning med blått / violett ljus ökar FMN-fotolys.
Detektion av  från FMN som bestrålas med blått eller violett ljus
från FMN som bestrålas med blått eller violett ljus
NBT-reduktion användes för att bestämma bildandet av 32. FMN är mycket känsligt för synligt och UV-ljus även under korta perioder av strålning. bildas från de mellanliggande fotokemiska reaktionerna under fotolysen av FMN. Figur 4 visar att den fotokemiska effekten av FMN på NBT-reduktion beror på bestrålningstiden, som bildas av FMN som utsätts för bestrålning av blått eller violett ljus på ett tidsberoende sätt. Den genomsnittliga fotolytiska effekten av blått ljus och violett ljus är dock liknande som visas i figur 4.
Effekt av FMN-fotolys på S. aureus livskraft
Effekten av FMN som utsätts för bestrålning av violett eller blått ljus på S. aureus livskraft bestämdes. FMN-fotolys med bestrålning av blått eller violett ljus resulterar i signifikant inaktivering av S. aureus. Ju större koncentration av FMN, desto högre inaktiveringsgrad för S. aureus som utsätts för bestrålning av blått ljus vid 20 W/m2 i 120 minuter enligt figur 5A. Inaktiveringsgrader på 26%, 39%, 43% och 97% uppnåddes för S. aureus med 0, 30, 60 respektive 120 μM FMN, under 20 W / m2 blåljusbestrålning i 120 min. Som visas i figur 5A finns det ingen signifikant skillnad (p-värde = 0,20) i inaktiveringsgraden för S. aureus med bestrålning av blått ljus med antingen 60 μM FMN vid 20 W/m 2 i 120 min (energidos, 14,4 J/cm 2) eller 120 μM FMN vid 20 W/m 2 i 60 min (energidos, 7,2 J/cm2). Å andra sidan, som visas i figur 5B, uppnåddes inaktiveringshastigheter på 53% och 96% för S. aureus utan respektive med 30 μM FMN under bestrålning av violett ljus vid 10 W/m 2 i 30 min (energidos på 1,8 J/cm2). Därför har FMN som utsätts för bestrålning av blått eller violett ljus en större effekt på livskraften hos S. aureus genom att öka bakterieinaktiveringen. FMN behandlad med bestrålning av violett ljus är effektivare vid S. aureus-inaktivering.
Detektion av i FMN-behandlad S. aureus under bestrålning av violett ljus
Produktionen av i FMN-behandlad S. aureus under bestrålning av violett ljus bestämdes med användning av NBT-reduktionsmetoden. Absorbansvärdet vid 560 nm användes för att utvärdera produktionens omfattning. Den fotokemiska effekten av FMN på generering i S. aureus är proportionell mot FMN-koncentrationen (figur 6). Absorbansvärdena på 560 nm för S. aureus utan FMN-behandling är 0,31 och 0,07 under 10 W/m2 bestrålning av violett ljus i 10 min respektive i mörker (figur 6). Detta tyder på att lite producerades i obehandlad S. aureus efter bestrålning av violett ljus. De respektive 560 nm absorbansvärdena för S. aureus behandlade med 100 μM FMN är 1,36 och 0,08 under 10 W/m2 violett ljusbestrålning i 10 min respektive i mörker. Därför ökar den fotokemiska effekten på NBT-reduktion i FMN-behandlad S. aureus under violett bestrålning, vilket produceras rikligt.

Figur 1: Schematisk representation av fotoreaktionssystemet. Klicka här för att se en större version av denna figur.

Figur 2: Emissionsspektra för blå, grön, röd och violett LED-lampor i denna studie. Emissionsmaxima för blå, grönt, rött och violett ljus är vid 465, 529, 632 respektive 405 nm, och spektralbredderna vid halvhöjd (W1/2) är 26, 31, 14 respektive 12 nm. Klicka här för att se en större version av denna siffra.

Figur 3: Absorbansspektra av FMN behandlade med lysdioder med olika våglängder vid 10 W/m2 i 5 min. Klicka här för att se en större version av denna figur.

Figur 4: NBT-reduktion för att bestämma effekten av bestrålning av blått och violett ljus vid 10 W/m2 i 1-5 min på FMN-fotolys. Klicka här för att se en större version av denna figur.

Figur 5: Effekt av FMN-fotolys på S. aureus livskraft. Den procentuella minskningen definieras som ett negativt värde för inaktiveringsgraden för S. aureus som behandlats med (A) FMN vid bestrålning av blått ljus vid 20 W/m 2 i 120 min och (B) FMN vid bestrålning av violett ljus vid 10 W/m2 i 30 min. FMN-koncentrationer och bestrålningstider anges högst upp i figuren. Observera att det inte finns någon signifikant skillnad (p = 0,20) i inaktiveringsgraden för S. aureus med 60 μM FMN vid 20 W/m 2 bestrålning av blått ljus i 120 min eller 120 μM FMN vid 20 W/m2 bestrålning av blått ljus i 60 min (A). Resultaten uttrycks som medelvärde ± SD, där n = 3. *p < 0,05; **p < 0,01; p < 0,0001. Klicka här för att se en större version av denna siffra.

Figur 6: Effekt av FMN-koncentration på generering i S. aureus utsatt för bestrålning av violett ljus vid 10 W/m2 i 10 min. Sammantaget produceras lite i S. aureus i frånvaro av FMN även om bestrålning av violett ljus ökar något i det bestrålade provet (PB (violett ljus)) i förhållande till mörk kontroll (PB (mörk)). Resultaten uttrycks som medelvärde ± SD, där n = 3. * p < 0,05; ** p < 0,01; p < 0,0001. Klicka här för att se en större version av denna siffra.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
En fotosensibiliserare ökar den fotokemiska reaktionen av kemiska föreningar för att generera ROS. Patogena mikroorganismer kan inaktiveras genom ljusbestrålning i närvaro av fotosensibiliserare. Denna studie bestämmer aPDI för S. aureus på grund av ROS genererad av bestrålning av violett ljus av en exogen fotosensibiliserare, FMN.
Som visas i figur 3, för FMN, reduceras absorbansen vid 444 nm signifikant efter 5 minuters bestrålning med violett eller blått ljus och det finns ingen signifikant förändring i FMN-absorbansen under bestrålning med grönt eller rött ljus. NBT-reduktion används för att bestämma bildandet av via en laddningsöverföringsprocess10,33. En tidigare studie av författarna använde FMN under blå, grön eller rött ljus bestrålning för att bestämma bildandet av att använda NBT-reduktion. Bestrålning av blått ljus ger en enorm mängd medan den fotolytiska effektiviteten hos grönt och rött ljus är mindre än 3% av det som uppnås med blått ljus10. Dessa resultat visar att en laddningsöverföringsprocess produceras när FMN utsätts för bestrålning av blått eller violett ljus och att det inte sker någon signifikant förändring vid bestrålning med grönt eller rött ljus.
En tidigare studie av författarna34 visade att mekanismen för FMN-fotolys kan beskrivas med primära FMN-fotolytiska reaktioner (ekvation 1):
 (1),
(1),
där 1FMN* och 3FMN* är singlet- respektive tripletttillstånden för det exciterade FMN:et. De fotolytiska reaktionsmekanismerna för dessa fotoinducerade FMN initieras möjligen av en triplet-triplet-förintelse eller en triplet-ground state quenching-process35,36,37,38. Grundtillståndet för FMN (FMN0) är en elektrondonator som ger en elektron till 3FMN* och genererar de halvreducerade (FMN•-) och halvoxiderade (FMN•+) formerna, som visas i ekvation 2
 (2)
(2)
Fotodegraderingsvägarna för FMN är resultatet av fotosensibilisering. FMN befordras till det elektroniskt exciterade tillståndet, FMN•, via en fotolytisk reaktion och sedan reduceras ett neutralt FMN när FMN• förlorar en elektron, som visas i ekvationerna 3-8 nedan:
 (3)
(3)
 (4)
(4)
 (5),
(5),
där FMN•+ är en radikal katjonart.
Samspelet mellan FMN•- medO2 (3Σg-) producerar den reaktiva superoxidradikalen, , följt av hydroperoxylradikalen, HOO•, som genereras i reaktionen mellan och en proton och så småningom leder till nedbrytning av FMN0:
 (6)
(6)
 (7)
(7)
 (8),
(8),
där FMN•- är en radikal anjon.
I den aktuella studien, i frånvaro av FMN, är inaktiveringsgraden för S. aureus 53% under bestrålning av violett ljus vid 10 W/m 2 i 30 min och 26% under bestrålning av blått ljus vid 20 W/m2 i 120 min, vilket tyder på att bestrålning av blått ljus inaktiverar färre bakterier än violett ljus (figur 5). Det har tidigare rapporterats att bestrålning vid våglängder längre än 430 nm utan en exogen fotosensibiliserare inte inaktiverar S. aureus-celler, även om porfyriner i S. aureus har en maximal absorbans vid 405 ± 5 nm och orsakar bakteriell inaktivering efter bestrålning39.
En tidigare studie av de nuvarande författarna visade att E. coli och S. aureus inaktiveras genom DNA-klyvning som induceras genom bildandet av genom fotolys av riboflavin eller FMN10,24,40. Respektive inaktiveringsgrad för S. aureus är 97 % respektive 96 % vid 120 μM FMN under 20 W/m 2 bestrålning av blått ljus (energidos på 14,4 J/cm 2) under 120 min och 30 μM FMN under 10 W/m 2 bestrålning av violett ljus (energidos på 1,8 J/cm 2) i 30 min (figur 5). Därför är bestrålning av violett ljus effektivare för att inaktivera S. aureus eftersom lågenergidoser av violett ljus kan inaktivera S. aureus i närvaro av lägre FMN-koncentrationer och kortare belysningsperioder jämfört med blått ljus. Även om omfattningen av FMN-fotolys som uppnås med violett ljus är högre jämfört med blått ljus (figur 3), är de genomsnittliga fotolytiska effekterna av FMN på NBT-reduktion under bestrålning av blått och violett ljus likartade (figur 4).
E. coli är en vanlig gramnegativ bakterie. Riboflavin eller FMN fotolys genom bestrålning av blått ljus inaktiverar E. coli via ROS 10,24,41 med en inaktiveringsgrad på 96% vid användning av FMN 10,24. S. aureus, å andra sidan, är en grampositiv bakterie, som effektivt inaktiveras av FMN-fotolys under bestrålning av violett ljus, vilket visas här (Figur 5). Därför kan FMN-fotolytisk reaktion under bestrålning av blått eller violett ljus producera ROS och inaktivera både gramnegativa och grampositiva bakterier.
I avsaknad av FMN produceras emellertid lite i S. aureus som utsätts för bestrålning av violett ljus (figur 6); därför minskar endogena fotosensibiliserare endast något livskraften hos S. aureus under bestrålning av violett ljus, jämfört med effekten i närvaro av exogen fotosensibiliserare såsom FMN. Under samma förhållanden minskar FMN som utsätts för bestrålning av violett ljus livskraften hos S. aureus för att resultera i en 96% inaktiveringsgrad, vilket är mycket högre än för endogena intracellulära fotosensibiliserare ensam. Produktionen av i FMN-behandlad S. aureus under bestrålning av violett ljus har ökat avsevärt, vilket visas i figur 6. Dessa resultat visar att FMN när det utsätts för bestrålning av violett ljus orsakar en förhöjd nivå av bakteriell inaktivering.
FMN kan uppnå ett fotoexciterat tillstånd vid bestrålning med UV 1,2,21,22,23, blå 10,24 och violett ljus 31; Den cytotoxicitet som orsakas av UV-strålning och högintensiv bestrålning av kortvåglängder måste dock beaktassom 42. Å andra sidan producerar röda och gröna strålningar med längre våglängder inte signifikanta mängder ROS under FMN-fotolys10. Därför kräver ROS-inducerad aPDI av S. aureus bestrålning av FMN med violett ljus som har en mellanliggande våglängd. Spektralbredden på halv höjd (W1/2) för violett ljus måste vara smal för att undvika överlappning med UV-absorptionen och hämma risken på grund av UV-strålning31. Mikronäringsämnet, FMN, är som sådant ofarligt och är ett viktigt vitamin för människokroppen. Med hjälp av en lämplig fotosensibiliserare som FMN kan bestrålning av violett ljus med låg energi undertrycka patogena bakterier, vilket ger ett säkert och effektivt medel för sanitetsprocessen. Förutom den fotolytiska reaktionen av FMN på S. aureus, förtjänar andra frågor ytterligare undersökning, såsom fototoxiciteten hos FMN på mikrober, som härrör från den stress som orsakas av , som kan studeras med hjälp av en RNA-seq-teknik tillsammans med qPCR.
FMN-fotolys via bestrålning av violett ljus leder till fotokemisk oxidation genom vilken ROS genereras och i sin tur ökar inaktiveringen av patogena bakterier. Användningen av violett ljus anses vara ett kritiskt steg i FMN-fotodekomposition. Begränsningen med den nuvarande tekniken är att våglängdsintervallet för violett ljus måste vara ganska smalt för att undvika risken på grund av UV-bestrålning. Den nuvarande metoden har potential för framtida medicinska tillämpningar, såsom infektionsbehandling av ythud tillsammans med djupa subkutana vävnader eller muskler genom att sätta in en optisk fiber för att rikta det violetta ljuset till de infekterade områdena. Fotokemisk behandling med FMN är därför ett säkert och enkelt sätt att säkerställa effektiva hygienrutiner.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna förklarar inga intressekonflikter.
Acknowledgments
Författarna är tacksamma mot Dr. Tak-Wah Wong och Mr. Zong-Jhe Hsieh för deras stöd med experiment.
Materials
| Name | Company | Catalog Number | Comments |
| Blue, green and red LED lights | Vita LED Technologies Co., Tainan, Taiwan | DC 12 V 5050 | |
| Dimethyl Sulfoxide | Sigma-Aldrich, St. Louis, MO | 190186 | |
| Infrared thermometer | Raytek Co. Santa Cruz, CA | MT4 | |
| LB broth | Difco Co., NJ | ||
| L-Methionine | Sigma-Aldrich, St. Louis, MO | 1.05707 | |
| NBT | Bio Basic, Inc. Markham, Ontario, Canada | ||
| Power supply | China tech Co., New Taipei City, Taiwan | YP30-3-2 | |
| Riboflavin 5′-phosphate | Sigma-Aldrich, St. Louis, MO | R7774 | |
| RNase | New England BioLabs, Inc. Ipswich, MA | ||
| Solar power meter | Tenmars Electronics Co., Taipei, Taiwan | TM-207 | |
| Staphylococcus aureus subsp. aureus | Bioresource Collection and Research Center (BCRC), Hsinchu, Taiwan | 10451 | |
| UV-Vis optical spectrometer | Ocean Optics, Dunedin, FL | USB4000 | |
| UV-Vis spectrophotometer | Hitachi High-Tech Science Corporation,Tokyo, Japan | U-2900 | |
| Violet LED | Long-hui Electronic Co., LTD, Dongguan, China |
References
- Jian, H. L., Cheng, C. W., Chen, L. Y., Liang, J. Y.
- Lin, Y., Eitenmiller, R. R., Landen, W. O. Riboflavin. Vitamin analysis for the health and food sciences. , CRC Press. 329-360 (2008).
- Xie, L. J., Wang, R. L., Wang, D., Liu, L., Cheng, L. Visible-light-mediated oxidative demethylation of N(6)-methyl adenines. Chemical Communications. 53 (77), 10734-10737 (2017).
- Tim, M. Strategies to optimize photosensitizers for photodynamic inactivation of bacteria. Journal of Photochemistry and Photobiology B: Biology. 150, 2-10 (2015).
- Maisch, T., et al. Photodynamic inactivation of multi-resistant bacteria (PIB) - a new approach to treat superficial infections in the 21st century. Journal of the German Society of Dermatology. 9 (5), 360-366 (2011).
- Hamblin, M. R. Antimicrobial photodynamic inactivation: a bright new technique to kill resistant microbes. Current Opinion in Microbiology. 33, 67-73 (2016).
- Del Rosso, J. Q. Oral Doxycycline in the management of acne vulgaris: Current perspectives on clinical use and recent findings with a new double-scored small tablet formulation. The Journal of Clinical and Aesthetic Dermatology. 8 (5), 19-26 (2015).
- Wong, T. W., et al. Photodynamic inactivation of methicillin-resistant Staphylococcus aureus by indocyanine green and near infrared light. Dematologica Sinica. 36 (1), 8-15 (2018).
- Yuann, J. M. P., et al. Effects of free radicals from doxycycline hyclate and minocycline hydrochloride under blue light irradiation on the deactivation of Staphylococcus aureus, including a methicillin-resistant strain. Journal of Photochemistry and Photobiology B: Biology. 226, 112370 (2022).
- Liang, J. Y., Cheng, C. W., Yu, C. H., Chen, L. Y. Investigations of blue light-induced reactive oxygen species from flavin mononucleotide on inactivation of E. coli. Journal of Photochemistry and Photobiology B: Biology. 143, 82-88 (2015).
- Yang, M. J., et al. Effects of blue-light-induced free radical formation from catechin hydrate on the inactivation of Acinetobacter baumannii, Including a carbapenem-resistant strain. Molecules. 23 (7), 1631 (2018).
- Yuann, J. M. P., et al. A study of catechin photostability using photolytic processing. Processes. 9 (2), 293 (2021).
- Yuann, J. M. P., Wang, J. S., Jian, H. L., Lin, C. C., Liang, J. Y. Effects of Clinacanthus nutans (Burm. f) Lindau leaf extracts on protection of plasmid DNA from riboflavin photoreaction. MC-Transaction on Biotechnology. 4 (1), 45-59 (2012).
- Rineh, A., et al. Attaching NorA efflux pump inhibitors to methylene blue enhances antimicrobial photodynamic inactivation of Escherichia coli and Acinetobacter baumannii in vitro and in vivo. Bioorganic and Medicinal Chemistry Letters. 28 (16), 2736-2740 (2018).
- Dai, T., et al. Photodynamic therapy for Acinetobacter baumannii burn infections in mice. Antimicrobial Agents and Chemotherapy. 53 (9), 3929-3934 (2009).
- Nitzan, Y., Ashkenazi, H. Photoinactivation of Acinetobacter baumannii and Escherichia coli B by a cationic hydrophilic porphyrin at various light wavelengths. Current Microbiology. 42 (6), 408-414 (2001).
- Tseng, C. C., et al. Altered susceptibility to the bactericidal effect of photocatalytic oxidation by TiO2 is related to colistin resistance development in Acinetobacter baumannii. Applied Microbiology and Biotechnology. 100 (19), 8549-8561 (2016).
- Boluki, E., et al. Antimicrobial activity of photodynamic therapy in combination with colistin against a pan-drug resistant Acinetobacter baumannii isolated from burn patient. Photodiagnosis and Photodynamic Therapy. 18, 1-5 (2017).
- Yang, M. Y., et al. Blue light irradiation triggers the antimicrobial potential of ZnO nanoparticles on drug-resistant Acinetobacter baumannii. Journal of Photochemistry and Photobiology B, Biology. 180, 235-242 (2018).
- Code of Federal Regulations. , Available from: https://www.gpo.gov/fdsys/pkg/CFR-2009-title21-vol6/pdf/CFR-2009-title21-vol6-sec582-5695.pdf (2022).
- Lu, C. Y., et al. Generation and photosensitization properties of the oxidized radical of riboflavin: a laser flash photolysis study. Journal of Photochemistry and Photobiology B, Biology. 52 (1-3), 111-116 (1999).
- Sato, K., et al. The primary cytotoxicity in ultraviolet-a-irradiated riboflavin solution is derived from hydrogen peroxide. The Journal of Investigative Dermatology. 105 (4), 608-612 (1995).
- Tripathi, A. K., et al. Attenuated neuroprotective effect of riboflavin under UV-B irradiation via miR-203/c-Jun signaling pathway in vivo and in vitro. Journal of Biomedical Science. 21 (1), 39 (2014).
- Liang, J. Y., et al. Blue light induced free radicals from riboflavin on E. coli DNA damage. Journal of Photochemistry and Photobiology B, Biology. 119, 60-64 (2013).
- Ottaway, P. B. The Technology of Vitamins in Food. , Chapman and Hall. London. Ch. 5 233-244 (1993).
- Kumar, V., et al. Riboflavin and UV-light based pathogen reduction: extent and consequence of DNA damage at the molecular level. Photochemistry and Photobiology. 80 (1), 15-21 (2004).
- Ito, K., Inoue, S., Yamamoto, K., Kawanishi, S. 8-Hydroxydeoxyguanosine formation at the 5'site of 5'-GG-3'sequences in double-stranded DNA by UV radiation with riboflavin. Journal of Biological Chemistry. 268 (18), 13221-13227 (1993).
- Liang, J. Y., Yuann, J. M. P., Hsie, Z. J., Huang, S. T., Chen, C. C. Blue light induced free radicals from riboflavin in degradation of crystal violet by microbial viability evaluation. Journal of Photochemistry and Photobiology B, Biology. 174, 355-363 (2017).
- Yang, M. Y., Chang, C. J., Chen, L. Y. Blue light induced reactive oxygen species from flavin mononucleotide and flavin adenine dinucleotide on lethality of HeLa cells. Journal of Photochemistry and Photobiology B, Biology. 173, 325-332 (2017).
- Cui, Z., Huang, Y., Mo, Q., Wang, X., Qian, K. Inactivation of lymphocytes in blood products using riboflavin photochemical treatment with visible light. Photochemistry and Photobiology. 84 (5), 1195-1200 (2008).
- Wong, T. W., Cheng, C. W., Hsieh, Z. J., Liang, J. Y. Effects of blue or violet light on the inactivation of Staphylococcus aureus by riboflavin-5′-phosphate photolysis. Journal of Photochemistry and Photobiology B: Biology. 173, 672-680 (2017).
- Russell, L. F., Vanderslice, J. T. Comprehensive review of vitamin B2 analytical methodology. Journal of Micronutrient Analysis. 8, 257-310 (1990).
- Cheng, C. W., Chen, L. Y., Chou, C. W., Liang, J. Y. Investigations of riboflavin photolysis via coloured light in the nitro blue tetrazolium assay for superoxide dismutase activity. Journal of Photochemistry and Photobiology B: Biology. 148, 262-267 (2015).
- Huang, S. T., et al. Effects of 462 nm light-emitting diode on the inactivation of Escherichia coli and a multidrug-resistant by tetracycline photoreaction. Journal of Clinical Medicine. 7 (9), 278 (2018).
- Barua, M. G., Escalada, J. P., Bregliani, M., Pajares, A., Criado, S. Antioxidant capacity of (+)-catechin visible-light photoirradiated in the presence of vitamin B2. Redox Report: Communications in Free Radical Research. 22 (6), 282-289 (2017).
- Castillo, C., Criado, S., Díaz, M., García, N. A. Riboflavin as a sensitiser in the photodegradation of tetracyclines. Kinetics, mechanism and microbiological implications. Dyes and Pigments. 72 (2), 178-184 (2007).
- Huvaere, K., Sinnaeve, B., Van Bocxlaer, J., Skibsted, L. H. Flavonoid deactivation of excited state flavins: reaction monitoring by mass spectrometry. Journal of Agricultural and Food Chemistry. 60 (36), 9261-9272 (2012).
- Massad, W. A., Bertolotti, S., Garcia, N. A. Kinetics and mechanism of the vitamin B2-sensitized photooxidation of isoproterenol. Photochemistry and Photobiology. 79 (5), 428-433 (2004).
- Maclean, M., MacGregor, S. J., Anderson, J. G., Woolsey, G. High-intensity narrow-spectrum light inactivation and wavelength sensitivity of Staphylococcus aureus. FEMS Microbiology Letters. 285 (2), 227-232 (2008).
- Huang, S. T., et al. The influence of the degradation of tetracycline by free radicals from riboflavin-5'-phosphate photolysis on microbial viability. Microorganisms. 7 (11), 500 (2019).
- Maisch, T., et al. Fast and effective photodynamic inactivation of multiresistant bacteria by cationic riboflavin derivatives. PLoS One. 9 (12), 111792 (2014).
- Grijzenhout, M., et al. Ultraviolet-B irradiation of platelets induces a dose-dependent increase in the expression of platelet activation markers with storage. British Journal of Haematology. 83 (4), 627-632 (1993).
