우리 환경에서 가장 유해 물질 중 하나 인 내분비 방해 화합물 (EDC)의 상당수가 있습니다. 이들은 주로 천연 자원에서 물을 오염 하는 에스트로겐 화합물. 그룹에 속하는 물질의 화학적 다양성은 검출을 위해 다른 분석 방법이 필요하기 때문에 존재에 대한 테스트를 어렵게 만듭니다. 그들의 화학 구조에 따라 물질이 실제로 에스트로겐으로 작용할 수 있는지 여부를 결정하는 것은 매우 어렵습니다. 또한, 이러한 물질은 환경에 있는 순수한 형태로 결코 존재하지 않습니다, 그래서 그들의 효력은 그밖 화합물에 의해 영향을 받을 수 있습니다, 너무1. 이러한 문제점은 에스트로겐 효과를 보이는 바이오모니터/바이오지표 유기체의 사용과 같은 효과검출 방법에 의해 해결될 수 있다2,,3,,4,,5.
최근에는 다양한 세포주6 및 효모 기반 시험 시스템2,,3이 에스트로겐성 효과를 검출하기 위해 개발되었다. 그러나, 이들은 일반적으로 에스트로겐 수용체2,,3에물질의 결합을 검출할 수 있다. 또한, 그들은 유기체에서 복잡 한 생리 과정을 모델링 할 수 없습니다., 또는 생활 단계의 호르몬 에 민감한 단계를 감지; 따라서, 그들은 종종 거짓 결과로 이어질.
특정 유전자가 살아있는 유기체7에서에스트로겐에 민감하게 반응하는 것으로 알려져 있다. 분자 생물학 방법에 의한 유전자 제품의 검출은 단백질 또는 mRNA 수준8,,9에서또한 가능하지만, 일반적으로 동물 희생을 수반한다. 동물보호법은 더욱 엄격해졌으며, 실험에 사용되는 동물의 수와 고통을 최소화하거나 동물 모델의 다른 모델시스템(10)을대체하는 대체 시험 시스템에 대한 수요가 증가하고 있다. 형광 단백질의 발견과 바이오 마커 라인의 생성과 함께, 형질 전환 기술은 좋은 대안11을제공합니다. 이 라인을 사용하여, 에스트로겐 에스트로겐 에민감한 유전자의 활성화는 생체 내에서 시험될 수 있습니다.
척추 동물 중, 환경 위험 평가에서 물고기의 잠재력은 뛰어난. 그들은 포유류 모델에 비해 많은 장점을 제공합니다 : 수생 유기체인, 그들은 몸 전체를 통해 오염 물질을 흡수 할 수 있습니다, 자손의 많은 수를 생산하고, 그들의 종의 일부는 짧은 세대 시간을 특징으로한다. 그들의 내분비 체계 및 생리 적 과정은 다른 척추 동물과 심지어 인간 을 포함한 포유 동물과 큰 유사성을 보여줍니다12.
물고기에 있는 에스트로겐 효과의 검출을 위한 몇몇 유전자는 또한 알려져 있습니다. 가장 중요한 것은 에스트로겐 수용체 아로마타제-b, 초리오게닌-H, 비텔로게닌(vtg)7,,13이다. 최근에는 제브라피쉬(Daniorerio)4,,5,,14,,15,,16, 17등 실험실에서 사용되는 생선 모델에서 여러 에스트로겐 생산 바이오센서 라인이 생성되고있다.17 바이오센서 라인을 만드는 데 있어 제브라피쉬의 주요 장점은 형광 기자 신호가동물(10)을희생시키지 않고 생체내에서 쉽게 연구될 수 있기 때문에 배아와 애벌레의 투명한 바디이다. 동물 보호 외에도, 치료18의다른 시기에 동일한 개인의 반응을 연구할 수 있기 때문에 귀중한 기능이기도 하다.
이러한 실험은 비텔로게닌 기자 트랜스제닉 제브라피쉬 라인15를사용합니다. Tg(vtg1:mCherry) 개발에 사용되는 트랜스진 구조는 긴(3.4kbp) 천연 비텔로게닌-1 프로모터를 갖는다. 에스트로겐 수용체(ER)는 스테로이드/핵 수용체 슈퍼패밀리를 대표하는 리간드에 의해 활성화된 증강 단백질이다. ER은 에스트로겐 반응 원소(ERE)라고 불리는 특정 DNA 서열에 높은 친화성을 가지며 에스트라디올 및 기타 에스트로겐 물질에 반응하여 유전자 발현을 트랜스활성화하므로 프로모터에서 ERE가 많을수록 더 강한 반응을일으킨다(19). Tg(vtg1:mCherry) 유전자 생성물의 프로모터 영역에는 17개의 ERE 부위가 있으며, 네이티브 vtg유전자(15)의발현을 모방할 것으로 예상된다. 성적으로 성숙한 여성에서 형광 신호의 지속적인 표현이 있다. 그러나, 남성 및 배아에서 간발현은 에스트로겐성 물질로 치료시만 볼 수있다(도 1).
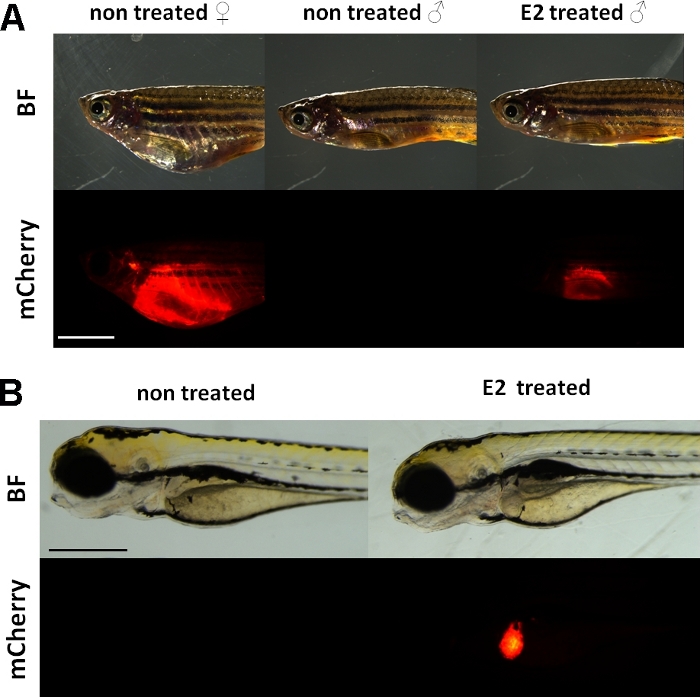
그림 1: vtg1:mCherry 트랜스제닉 성 제브라피시와 5dpf 배아의 간에서 적색 형광 신호, 17-ß-estradiol (E2) 유도에 따라. 여성 및 E2(25 μg/L 노출 시간:48시간:48시간)로 치료된 남성에서는 간의 강한 형광이 안료 피부를 통해서도 볼 수 있다. 치료되지 않은남성(A)에서형광 신호가 보이지 않습니다. E2 유도(50 μg/L 노출 시간: 0-120 hpf)에 이어, 5dpf 배아의 간에서 적색 형광 신호도 관찰될B수 있으며, 이는 대조군 배아(B)에서 볼 수 없다. 형광 신호는 성인 여성에서 지속적으로 존재하는 동안, 주로 남성과 라인의 배아는 에스트로겐 효과를 검출하기에 적합합니다. (BF: 밝은 필드, mCherry: 빨간 형광 필터 보기, 단일 일반 이미지, 스케일 바 A: 5mm, 스케일 바 B: 250 μm) 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
내인성 비텔로게닌과 마찬가지로, mCherry 리포터는 간에서만 표현됩니다. 비텔로게닌은 에스트로겐의 존재에서만 생산되기 때문에 컨트롤에는 형광 신호가 없습니다. 발현은 간에서만 있기 때문에 결과의 평가는훨씬 쉽게 15.
이 선의 배아의 민감도 및 유용성은 다양한 에스트로겐 화합물 혼합물과 또한 환경 샘플15,,20에대해 조사되었으며, 대부분의 경우 용량 반응 관계가 문서화되었다(그림2). 그러나, 고독성, 주로 간독성, 물질(예를 들어, 제랄레네네)의 경우, 처리된 배아의 간에서 매우 약한 형광 신호만이 볼 수 있으며, 발생되는 최대 강도형 형광 신호는 매우 작은 농도 범위 내에 도달할 수 있어 투여 효과관계(20)를확립하기 어렵게 만든다.
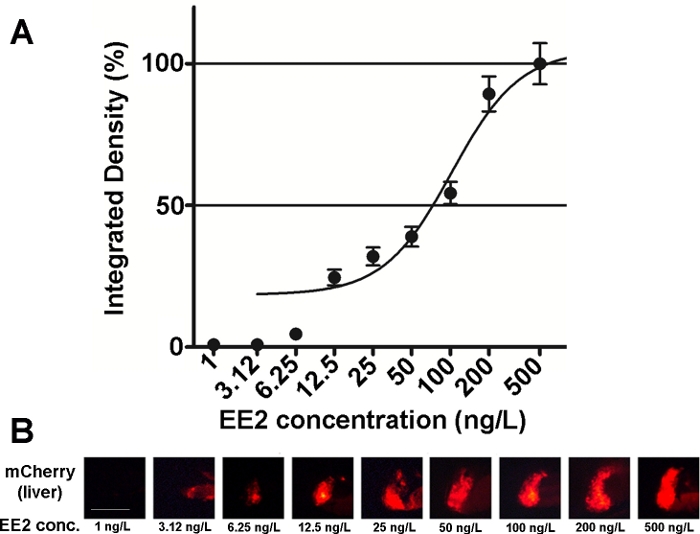
도 2: 투여 반응 다이어그램(A) 및 형광 이미지(mCherry)가 17-α-ethynilestradiol(EE2)에 노출된 간(B)의 형광 이미지(mCherry)를 5dpf vtg1:mCherry 애벌레로 노출시켰다. 결과는 신호 강도 및 영향 영역의 크기(±SEM, n = 60)로부터 생성된 통합 밀도로 표현됩니다. 100% 관찰된 최대값을 나타냅니다. 형광 신호 강도는 농도로 점차적으로 증가했습니다. 스케일 바 = 250 μm. 여기를 클릭하여 이 그림의 더 큰 버전을 확인하십시오.
환경에는 몇 가지 에스트로겐 성 물질이 존재합니다. 17-β-estradiol (환경 농도: 0.1-5.1 ng/L)21,17-α-에티닐에스트라디오(환경 농도: 0.16-0.2 μg/L)2 2,제랄레네(환경 농도: 0.095-0.22 μg/L)23,비스페놀-A(환경 농도: 0.45-17.2 mg/L)24. mCherry 형질전환 배아의 도움으로 순수한 활성 형태로 이러한 물질을 테스트 할 때, 형광 기호 검출을 위한 가장 낮은 관찰 효과 농도(LOEC)는 17-ß-estradiol용 100 ng/L, 17-α-에티닐에스트라디오l용 1ng/L, 제아라레놀용 100ng/L, 비스페놀-A(96-120hpf)15내1mg/L, 1mg/L 또는 1/1의 근사 또는 환경범위 내에 매우 근접한 15개의 환경범위 내에 있었다. Tg (vtg1:mCherry) 형질 전환선은 직접 노출 후 폐수 샘플에서 에스트로겐성을 감지하는 데 도움이 될 수 있습니다. 라인은 일반적으로 사용되는 효모 에스트로겐 시험, 생물루미니스센트 효모 에스트로겐(BLYES) 분석15만큼민감하다. 이 라인의 도움으로, 제랄레네유발 독성에 대한 베타-사이클로덱스트린의 보호 효과는 화학혼합물(20)을사용하여 확인되었다.
최근 보고서에서, 형질대사의 생체 내 사용은 2개의 에스트로겐제일릭 제아롤론(ZEA) 대사산물, α-및 β-제아롤레놀(α-ZOL 및 β-ZOL)25의도움으로 입증되었다. 프로토콜 기준선은 Tg (vtg1:mCherry) 배아에 여러 화합물 또는 환경 샘플의 에스트로겐 효과를 연구하는 것이 적절하다.
