在我们的环境中,有大量内分泌干扰化合物(EDC)是我们环境中最危险的物质之一。这些主要是从自然资源中污染水的雌激素化合物。属于该组的物质的化学多样性使得测试其存在变得困难,因为检测这些物质需要不同的分析方法。根据它们的化学结构,很难确定一种物质是否真的能够充当雌激素。此外,这些物质永远不会以纯形式存在于环境中,因此它们的影响可能受其他化合物的影响,太1。这个问题可以通过效果检测方法解决,例如使用生物监测器/生物致病生物,显示雌激素效应2,3,4,5。3,4,52
最近,各种细胞系6和酵母为基础的测试系统22,33已经开发出来检测雌激素效应。然而,这些通常只能检测物质与雌激素受体2,3,的结合。此外,它们无法模拟生物体中复杂的生理过程,或检测生命阶段的激素敏感阶段;因此,它们往往导致错误的结果。
众所周知,某些基因对生物体中的雌激素反应敏感。通过分子生物学方法检测基因产品也可以在蛋白质或mRNA级别8,9,9但通常涉及动物牺牲。8动物保护法已经变得更加严格,对替代性测试系统的需求也日益增长,这种试验系统可以最大限度地减少实验中使用的动物的数量和痛苦,或者用另一个模型系统10取代动物模型。随着荧光蛋白的发现和生物标志物线的产生,转基因技术提供了很好的替代方案。有了这些线,雌激素敏感基因的激活可以在体内测试。
在脊椎动物中,鱼类在环境风险评估中的潜力是突出的。与哺乳动物模型相比,它们有许多优点:作为水生生物,它们能够通过整个身体吸收污染物,产生大量的后代,而且它们的一些物种的特点是生成时间短。它们的内分泌系统和生理过程与其他脊椎动物,甚至哺乳动物,包括人类12有很大的相似之处。
几种用于检测鱼类雌激素效应的基因也已知。最重要的是雌激素受体芳香酶-b,胆碱-H,和维泰洛宁(vtg)7,13。7,13最近,一些产生雌激素的生物传感器线也创造了从鱼模型在实验室使用,如从斑马鱼(达尼奥雷里奥)4,5,14,15,16,17。4,5,14,15,16,17斑马鱼在创建生物传感器线的主要优点是胚胎和幼虫的透明体,因为荧光报告器信号可以在体内轻松研究,而不会牺牲动物10。除了动物保护,它也是一个有价值的功能,因为它允许研究同一个人的反应在治疗的不同时间18。
这些实验使用维泰洛根因记者转基因斑马鱼线15。用于开发Tg(vtg1:mCherry)的转基因结构具有长(3.4 kbp)天然维泰洛根-1启动子。雌激素受体(ER)是由配体激活的增强蛋白,是类固醇/核受体超级家庭的代表。ER结合到特定的DNA序列称为雌激素反应元素(EREs),具有高亲和力和转活性基因表达对雌二醇和其他雌激素物质的反应,所以在启动子中更多的ERE导致更强的反应19。在Tg(vtg1:mCherry)的启动区域有17个ERE位点,它们有望模仿原生vtg基因15的表达。在性成熟的女性中,荧光信号有连续的表达。然而,在雄性和胚胎中,肝脏中的表达只有在雌激素治疗时才能看到(图1)。
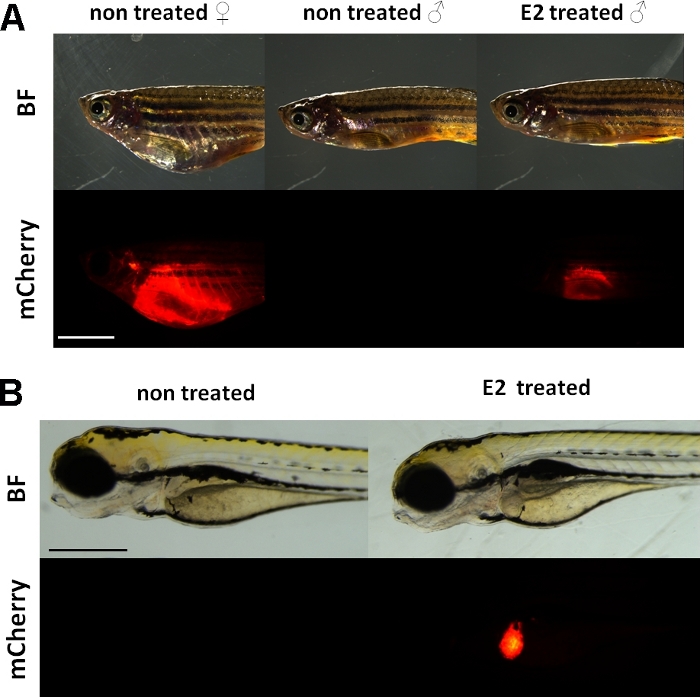
图1:vtg1:mCherry转基因成年斑马鱼和5dpf胚胎肝脏的红色荧光信号,17-β-雌二醇(E2)诱导后。在女性和男性治疗E2(25μg/L暴露时间:48小时)肝脏强荧光是可见的,即使通过色素的皮肤。未经治疗的雄性(A)看不到荧光信号。E2 诱导(50 μg/L 暴露时间:0-120 hpf)后,还可以观察到 5 dpf 胚胎肝脏中的红色荧光信号,这在控制胚胎(B) 中不可见。虽然荧光信号持续存在于成年女性体内,但主要为男性和胚胎,适合检测雌激素效应。(BF:亮场,mCherry:红色荧光滤光片视图,单纯图像,比例杆 A:5mm,比例杆B:250μm)请单击此处查看此图的较大版本。
与内源性维泰洛源类似,mCherry 记者只在肝脏中表达。由于维泰洛根宁只在雌激素存在的情况下产生,因此控制装置中没有荧光信号。因为表达只在肝脏,评估结果要容易得多15。
该线的胚胎的敏感性和可用性已研究的各种雌激素化合物混合物,以及环境样本15,20,在大多数情况下剂量反应15,20关系被记录(图2)。然而,在剧毒,主要是肝毒性,物质(如泽拉酮),只有非常弱的荧光信号可能可见于治疗胚胎的肝脏,最大强度荧光信号可以达到一个非常小的浓度范围内,这使得很难建立剂量效应关系20。
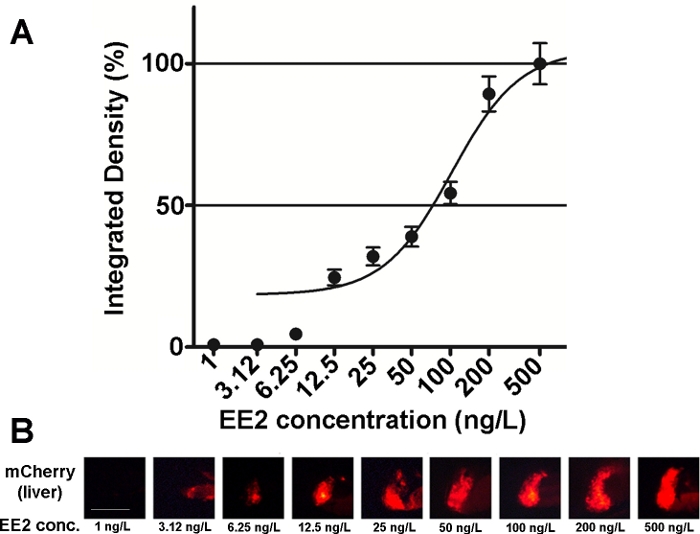
图2:肝脏的剂量反应图(A)和荧光图像(mCherry),在5dpf vtg1:mCherry幼虫中,暴露于17-β-乙二醇二醇(EE2)。结果表示为从信号强度和受影响区域大小(+SEM,n = 60)生成的集成密度。100% 是指观测到的最大值。荧光信号强度随着浓度的逐渐增加。比例线 = 250 μm. 请单击此处查看此图的较大版本。
环境中存在几种雌激素物质, 如17-μ-雌二醇(环境浓度:0.1±5.1纳克/升)21,17-β-乙酰二醇(环境浓度:0.16~0.2μg/L)22,泽拉酮 (环境浓度: 0.095~0.22 μg/L)23, 双酚-A (环境浓度: 0.45~17.2毫克/升)24.当在转基因胚胎的帮助下以纯活性形式测试这些物质时,用于荧光标志检测的最低观测效果浓度(LOEC)为17-β–雌二醇的100纳克/升, 1 ng/L 代表 17-α-乙酰二醇,100 ng/L 代表泽拉酮,1 mg/L 代表双酚-A(96-120 hpf 处理),非常接近或非常接近15.的环境浓度范围。Tg(vtg1:mCherry)转基因线有助于在直接接触后检测废水样品中的雌激素。线是敏感作为常用酵母雌激素测试,生物卢米尼森酵母雌激素(BLYES)测定15。在这条线的帮助下,使用化学混合物20证实了β-环糊精对泽拉酮诱导毒性的保护作用。
在最近的一份报告中,在两种雌激素泽拉酮(ZEA)代谢物的帮助下,在体内使用转基因线,α-泽拉醇(+-ZOL和+-ZOL)25。协议基线适合研究几种化合物或环境样本对Tg(vtg1:mCherry)胚胎的雌激素效应。
