Es gibt eine erhebliche Anzahl von endokrin störenden Verbindungen (EDC), die zu den gefährlichsten Stoffen in unserer Umwelt gehören. Dies sind vor allem östrogene Verbindungen, die Wasser aus natürlichen Ressourcen kontaminieren. Die chemische Vielfalt der zur Gruppe gehörenden Stoffe erschwert die Prüfung auf ihre Anwesenheit, da für ihren Nachweis unterschiedliche Analysemethoden erforderlich sind. Aufgrund ihrer chemischen Struktur ist es sehr schwierig festzustellen, ob ein Stoff tatsächlich in der Lage ist, als Östrogen zu wirken. Darüber hinaus sind diese Stoffe nie in reiner Form in der Umwelt vorhanden, so dass ihre Auswirkungen auch durch andere Verbindungen beeinflusst werden können, auch1. Dieses Problem kann durch wirkungserkennende Methoden gelöst werden, wie die Verwendung von Biomonitor-/Bioindikatororganismen, die östrogene Effekte zeigen2,3,4,5.
Kürzlich wurden eine Vielzahl von Zelllinie6 und Hefe-basierte Testsysteme2,3 entwickelt, um östrogene Effekte zu erkennen. Diese sind jedoch in der Regel nur in der Lage, die Bindung der Substanz an den Östrogenrezeptor2,3zu erkennen. Darüber hinaus sind sie nicht in der Lage, komplexe physiologische Prozesse im Organismus zu modellieren oder hormonempfindliche Phasen von Lebensphasen zu erkennen; daher führen sie oft zu falschen Ergebnissen.
Es ist bekannt, dass bestimmte Gene empfindlich auf Östrogen in lebenden Organismen reagieren7. Der Nachweis von Genprodukten durch molekularbiologische Methoden ist auch auf der Protein- oder mRNA-Ebene8,9möglich, beinhaltet aber in der Regel Tieropfer. Die Tierschutzgesetze sind strenger geworden, und die Nachfrage nach alternativen Testsystemen, die die Anzahl und das Leiden der in Experimenten verwendeten Tiere oder die Ersetzung des Tiermodells durch ein anderes Modellsystem minimieren, ist gestiegen10. Mit der Entdeckung fluoreszierender Proteine und der Schaffung von Biomarker-Linien bieten transgene Technologien eine gute Alternative11. Mit diesen Linien kann die Aktivierung eines Östrogen-sensitiven Gens in vivo getestet werden.
Bei Wirbeltieren ist das Potenzial von Fischen bei der Umweltverträglichkeitsprüfung hervorragend. Sie bieten viele Vorteile gegenüber Säugetiermodellen: Als Wasserorganismen sind sie in der Lage, Schadstoffe durch ihren gesamten Körper zu absorbieren, produzieren eine große Anzahl von Nachkommen, und einige ihrer Arten zeichnen sich durch kurze Erzeugungszeit aus. Ihr endokrines System und physiologische Prozesse zeigen große Ähnlichkeiten mit anderen Wirbeltieren und sogar mit Säugetieren, einschließlich Menschen12.
Mehrere Gene zum Nachweis von östrogene Wirkungen bei Fischen sind ebenfalls bekannt. Die wichtigsten sind die Östrogen-Rezeptoren Aromatase-b, Choriogenin-H, und Vitellogenin (vtg)7,13. Kürzlich wurden mehrere Östrogen-produzierende Biosensorlinien auch aus Fischmodellen erstellt, die im Labor verwendet werden, wie z.B. von Zebrafischen (Danio rerio)4,5,14,15,16,17. Der Hauptvorteil von Zebrafischen bei der Erstellung von Biosensorlinien ist der transparente Körper der Embryonen und Larven, da das fluoreszierende Reportersignal dann einfach in vivo untersucht werden kann, ohne das Tier10zu opfern. Neben dem Tierschutz ist es auch ein wertvolles Merkmal, da es ermöglicht, die Reaktion des gleichen Individuums zu verschiedenen Zeiten der Behandlung zu untersuchen18.
Diese Experimente verwenden einen Vitellogenin Reporter transgene Zebrafischlinie15. Das transgene Konstrukt, das für die Entwicklung von Tg(vtg1:mCherry) verwendet wird, hat einen langen (3,4 kbp) natürlichen Vitellogenin-1-Promotor. Der Östrogen-Rezeptor (ER) ist ein Enhancer-Protein durch Liganden aktiviert, die ein Vertreter der Steroid / Kernrezeptor Superfamilie ist. ER bindet an spezifische DNA-Sequenzen, sogenannte Östrogen-Response-Elemente (EREs) mit hoher Affinität und transaktiviert die Genexpression als Reaktion auf Ösradiol und andere östrogene Substanzen, so dass je mehr ERE im Promotor eine stärkere Reaktion verursacht19. Es gibt 17 ERE-Standorte in der Promotorregion des Tg(vtg1:mCherry) Transgenkonstrukts und es wird erwartet, dass sie die Expression des nativen vtg-Gens15imitieren. Es gibt einen kontinuierlichen Ausdruck des fluoreszierenden Signals bei geschlechtsreifen Weibchen. Bei Männern und Embryonen ist die Expression in der Leber jedoch nur bei der Behandlung mit östrogenen Substanzen sichtbar (Abbildung 1).
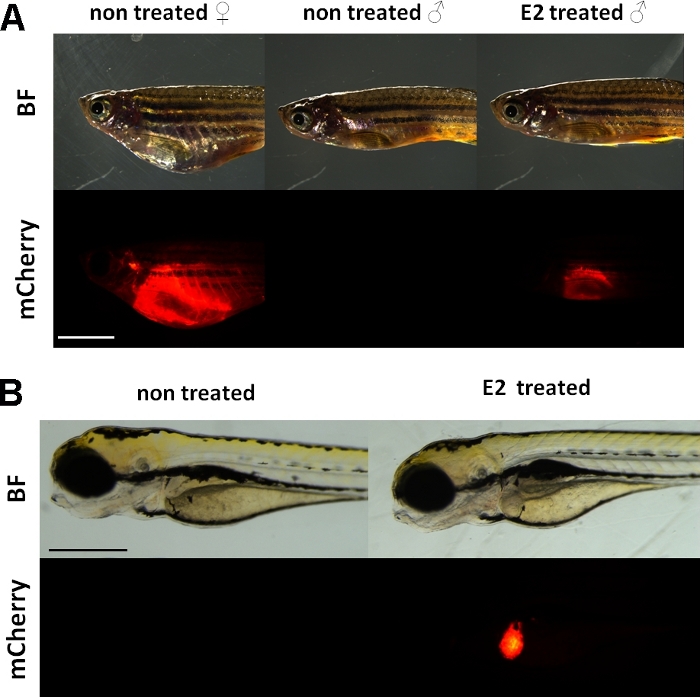
Abbildung 1: Rotes Fluoreszenzsignal in der Leber von vtg1:mCherry transgene erwachsene Zebrafische und 5 dpf Embryonen, nach 17-ß-Estradiol (E2) Induktion. Bei Frauen und Männern, die mit E2 (25 g/L Expositionszeit:48 Stunden) behandelt werden, ist eine starke Fluoreszenz der Leber auch durch die pigmentierte Haut sichtbar. Bei unbehandeltem Männchen ist kein fluoreszierendes Signal sichtbar (A). Nach der E2-Induktion (50 g/L-Expositionszeit: 0-120 hpf) kann auch ein rotes fluoreszierendes Signal in der Leber von 5 dpf-Embryonen beobachtet werden, das in Kontrollembryonen nicht sichtbar ist (B). Während das fluoreszierende Signal kontinuierlich bei erwachsenen Weibchen vorhanden ist, eignen sich vor allem Männchen und Embryonen der Linie zum Nachweis östrogener Wirkungen. (BF: helles Feld, mCherry: rote fluoreszierende Filteransicht, einzelne einfache Bilder, Skalenleiste A: 5mm, Skalenbalken B: 250 m) Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Ähnlich wie das endogene Vitellogenin wird der mCherry-Reporter nur in der Leber ausgedrückt. Da Vitellogenin nur in Gegenwart von Östrogen produziert wird, gibt es kein fluoreszierendes Signal in den Kontrollen. Da die Expression nur in der Leber ist, ist die Auswertung der Ergebnisse viel einfacher15.
Die Empfindlichkeit und Verwendbarkeit der Embryonen dieser Linie wurde an verschiedenen östrogenen Mischungen und auch an Umweltproben15,20, und in den meisten Fällen Dosis-Wirkungs-Beziehungen dokumentiert (Abbildung 2). Bei hochgiftigen, hauptsächlich hepatotoxischen Substanzen (z.B. Zearalenon) kann jedoch nur ein sehr schwaches fluoreszierendes Signal in der Leber behandelter Embryonen sichtbar sein und das verursachte maximale Fluoreszenzsignal kann innerhalb eines sehr kleinen Konzentrationsbereichs erreicht werden, was es schwierig macht, Dosis-Wirkungs-Beziehungen zu etablieren20.
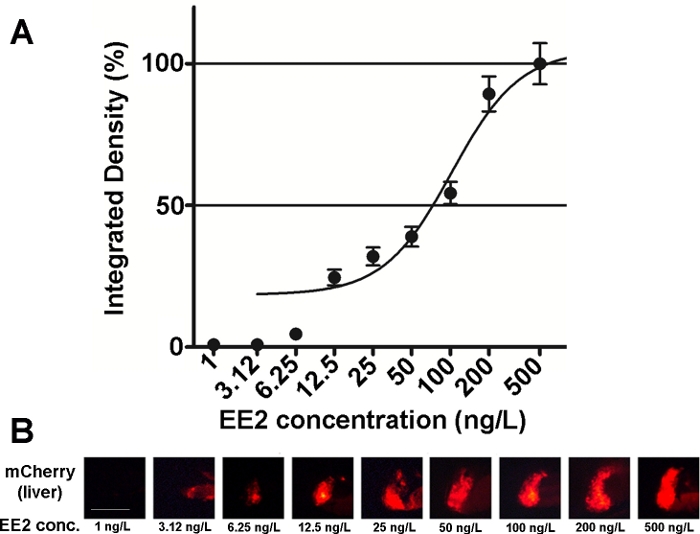
Abbildung 2: Dosis-Wirkungs-Diagramm (A) und fluoreszierende Bilder (mCherry) der Leber (B), die 17-A-Ethynilestradiol (EE2) ausgesetzt sind, in 5 dpf vtg1:mCherry Larven. Die Ergebnisse werden als integrierte Dichte ausgedrückt, die aus der Signalstärke und der Größe des betroffenen Bereichs erzeugt wird (SEM, n = 60). 100% bezieht sich auf das beobachtete Maximum. Die fluoreszierende Signalintensität nahm mit der Konzentration allmählich zu. Maßstabsleiste = 250 m. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Es gibt mehrere östrogene Substanzen in der Umwelt, wie z. B. 17-‘-Estradiol (Umweltkonzentration: 0,1–5,1 ng/L)21, 17-A-Ethynylestradiol (Umweltkonzentration: 0,16–0,2 g/L))22, Zearalenon (Umweltkonzentration: 0,095–0,22 g/L)23, Bisphenol-A (Umweltkonzentration: 0,45–17,2 mg/L)24. Bei der Prüfung dieser Stoffe in rein aktiver Form mit Hilfe von mCherry transgenen Embryonen die niedrigsten beobachteten Wirkungskonzentrationen (LOEC) für den Fluoreszenzzeichennachweis waren 100 ng/L für 17-ß-Estradiol, 1 ng/L für 17-A-Ethynilestradiol, 100 ng/L für Zearalenon und 1 mg/L für Bisphenol-A (96–120 h-Behandlung), die sehr nahe oder innerhalb des Bereichs der Umweltkonzentrationen der Stoffe15liegt. Die transgene Tg(vtg1:mCherry) kann helfen, Östrogeneität in Abwasserproben nach direkter Exposition zu erkennen. Die Linie ist so empfindlich wie der häufig verwendete Hefe-Östrogen-Test, der bioluminiscent Hefe-Östrogen (BLYES) Assay15. Mit Hilfe dieser Linie wurde die schützende Wirkung von Beta-Cyclodextrinen gegen zearalenon-induzierte Toxizität mit chemischen Gemischen bestätigt20.
In einem kürzlich veröffentlichten Bericht wurde die In-vivo-Verwendung der transgenen Linie mit Hilfe von zwei östrogenenen Zearalenon (ZEA)-Metaboliten, den Metaboliten , und -zearalenol (-ZOL und -ZOL)25, nachgewiesen. Die Protokollbasis ist geeignet, die östrogene Wirkung mehrerer Verbindungen oder Umweltproben auf Tg(vtg1:mCherry)-Embryonen zu untersuchen.
