Det finns ett betydande antal hormonstörande föreningar (EDC) som är bland de farligaste ämnena i vår miljö. Dessa är främst östrogena föreningar som förorenar vatten från naturresurser. Den kemiska mångfalden av de ämnen som tillhör gruppen gör det svårt att testa deras närvaro, eftersom det krävs olika analysmetoder för att de ska kunna upptäckas. Baserat på deras kemiska struktur är det mycket svårt att avgöra om ett ämne faktiskt kan fungera som ett östrogen. Dessutom förekommer dessa ämnen aldrig i ren form i miljön, så deras effekter kan påverkas av andra föreningar, för1. Detta problem kan lösas genom effekt-upptäcka metoder, såsom användning av biomonitor/ bioindicator organismer som visar östrogena effekter2,3,4,5.
Nyligen, en mängd celllinje6 och jäst-baserade testsystem2,3 har utvecklats för att upptäcka östrogena effekter. Emellertid, dessa är i allmänhet endast kunna upptäcka bindning av ämnet till östrogenreceptorn2,3. Dessutom kan de inte modellera komplexa fysiologiska processer i organismen, eller att upptäcka hormonkänsliga faser av livsstadier; därför leder de ofta till falska resultat.
Det är känt att vissa gener reagerar känsligt på östrogen i levande organismer7. Detektion av genprodukter genom molekylärbiologiska metoder är också möjligt på protein- eller mRNA-nivå8,9, men innebär vanligtvis djuroffer. Djurskyddslagar har blivit strängare, och det finns en växande efterfrågan på alternativa testsystem som minimerar antalet och lidandet hos djur som används i experiment eller ersätter djurmodellen med ett annat modellsystem10. Med upptäckten av fluorescerande proteiner och skapandet av biomarkörlinjer ger transgen teknik ett bra alternativ11. Med dessa linjer, aktivering av en östrogen-känslig gen kan testas in vivo.
Bland ryggradsdjur är fiskens potential i miljöriskbedömningen enastående. De erbjuder många fördelar jämfört med däggdjursmodeller: att vara vattenlevande organismer, de kan absorbera föroreningar genom hela kroppen, producera ett stort antal avkommor, och några av deras arter kännetecknas av kort generationstid. Deras endokrina system och fysiologiska processer visar stora likheter med andra ryggradsdjur och även med däggdjur, inklusive människor12.
Flera gener för att upptäcka östrogena effekter hos fisk är också kända. De viktigaste är östrogenreceptorerna aromatas-b, choriogenin-H och vitellogenin (vtg)7,13. Nyligen har flera östrogenproducerande biosensorlinjer också skapats från fiskmodeller som används i laboratoriet, till exempel från zebrafiskar (Danio rerio)4,5,14,15,16,17. Den största fördelen med zebrafiskar i att skapa biosensor linjer är den genomskinliga kroppen av embryon och larver, eftersom fluorescerande reporter signal kan sedan lätt studeras in vivo utan att offra djuret10. Förutom djurskydd är det också en värdefull funktion eftersom det gör det möjligt att studera reaktionen hos samma individ vid olika tidpunkter av behandlingen18.
Dessa experiment använder en vitellogenin reporter transgena zebrafisk linje15. Transgenen konstruktion som används för utveckling av Tg (vtg1:mCherry) har en lång (3,4 kbp) naturlig vitellogenin-1 promotor. Östrogenreceptorn (ER) är ett förstärkare protein som aktiveras av ligander som är en representant för steroid/nukleära receptorn superfamilj. ER binder till specifika DNA-sekvenser som kallas östrogen svarselement (EREs) med hög affinitet och transaktiverar genuttryck som svar på östradiol och andra östrogena ämnen, så ju mer ERE i promotorn orsakar ett starkare svar19. Det finns 17 ERE platser i promotorn regionen Tg (vtg1:mCherry) transgen konstruktion och de förväntas efterlikna uttrycket av den infödda vtggenen 15. Det finns ett kontinuerligt uttryck för fluorescerande signal hos sexuellt mognade kvinnor. Hos män och embryon syns dock uttrycket i levern endast vid behandling med östrogena ämnen (figur 1).
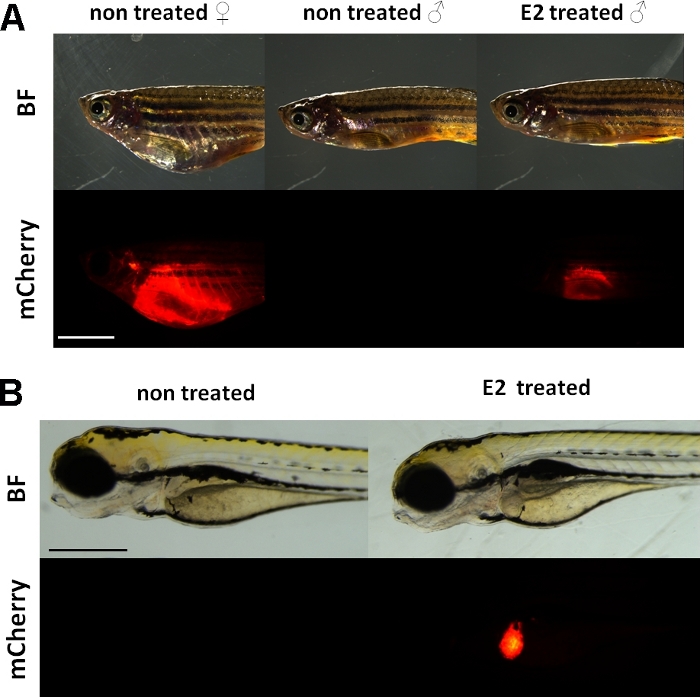
Figur 1: Röd fluorescerande signal i levern av vtg1:mRenasgen vuxen zebrafisk och 5 dpf embryon, efter 17-ß-estradiol (E2) induktion. Hos hona och hos hanar som behandlats med E2 (25 μg/L exponeringstid:48 timmar) syns stark fluorescens i levern även genom den pigmenterade huden. Ingen lysrörssignal syns hos obehandlad hane (A). Efter E2 induktion (50 μg/L exponeringstid: 0-120 hpf) kan en röd fluorescerande signal i levern på 5 dpf-embryon också observeras, vilket inte syns i kontrollembryon (B). Medan fluorescerande signal är kontinuerligt närvarande hos vuxna kvinnor, främst män och embryon i linjen är lämpliga för att upptäcka östrogena effekter. (BF: ljust fält, mCherry: röd lysrörsfiltervy, enkla vanliga bilder, Skalfält A: 5mm, skalstång B: 250 μm) Klicka här för att se en större version av denna siffra.
I likhet med den endogena vitellogenin uttrycks mCherry-reportern endast i levern. Eftersom vitellogenin endast produceras i närvaro av östrogen, det finns ingen fluorescerande signal i kontrollerna. Eftersom uttrycket är bara i levern, utvärderingen av resultaten är mycket lättare15.
Känsligheten och användbarheten hos denna linjes embryon har undersökts på olika östrogena föreningsblandningar och även på miljöprover15,,20,och i de flesta fall dokumenterades dos-responsförhållanden (figur 2). När det gäller mycket giftiga, huvudsakligen hepatotoxiska, ämnen (t.ex. iver) får dock endast en mycket svag lysrörssignal vara synlig i levern hos behandlade embryon och den maximala lysrörssignal som orsakas kan uppnås inom ett mycket litet koncentrationsområde, vilket gör det svårt att fastställa doseffektförhållanden20.
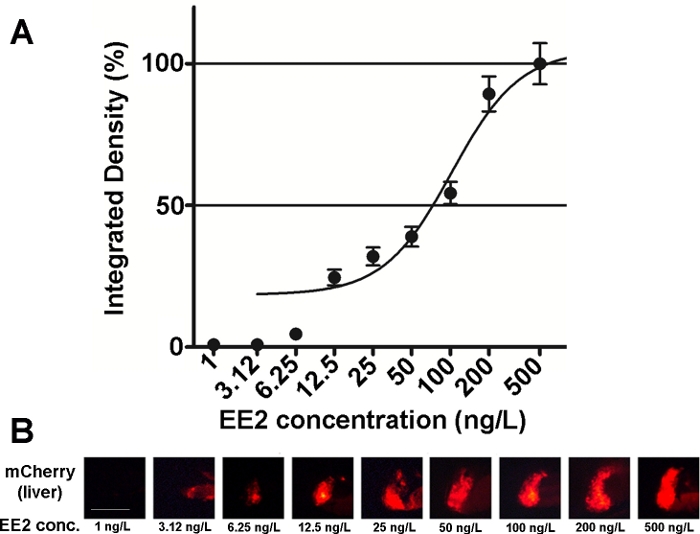
Figur 2: Dosresponsdiagram (A) och fluorescerande bilder (mCherry) i levern (B) som utsätts för 17-α-ethynilestradiol (EE2), i 5 dpf vtg1:mCherry larver. Resultaten uttrycks som integrerad densitet som genereras från signalstyrkan och storleken på det drabbade området (±SEM, n = 60). 100% avser det observerade maximivärdet. Fluorescerande signalintensitet ökade gradvis med koncentration. Skala bar = 250 μm. Klicka här för att se en större version av denna siffra.
Det finns flera östrogena ämnen som finns i miljön, såsom 17–β-estradiol (miljökoncentration: 0,1–5,1 ng/L)21,17–α-etynylestradiol (miljökoncentration: 0,16–0,2 μg/L)22, iveradon (miljökoncentration: 0,095–0,22 μg/L)23, bisfenol-A (miljökoncentration: 0,45–17,2 mg/L)24. Vid testning av dessa ämnen i ren aktiv form med hjälp av mCherry-transgena embryon var de lägsta koncentrationerna av observerade effekter (LOEC) för fluorescerande teckendetektering 100 ng/L för 17-ß-estradiol. 1 ng/L för 17-α-ethynilestradiol, 100 ng/L för iardon och 1 mg/L för bisfenol-A (behandling med 96–120 hpf), som ligger mycket nära eller inom intervallet för miljökoncentrationerna av ämnena15. Den Tg (vtg1:mCherry) transgen linje kan hjälpa till att upptäcka östrogenitet i avloppsvatten prover efter direkt exponering. Linjen är lika känslig som den vanliga jäst östrogen test, den bioluminiscent jäst östrogen (BLYES) analys15. Med hjälp av denna linje har de skyddande effekterna av beta-cyklodextriner mot iaralenoninducerad toxicitet bekräftats med hjälp av kemiska blandningar20.
I en färsk rapport demonstrerades in vivo-användningen av den transgena linjen med hjälp av två östrogena zearalenon (ZEA) metaboliter, α- och β-zearalenol (α-ZOL och β-ZOL)25. Protokollet baslinjen är lämplig att studera de östrogena effekterna av flera föreningar eller miljöprover på Tg (vtg1:mCherry) embryon.
