De modulatie van neuronale circuits in het centrale zenuwstelsel en hun gedragsresultaten zijn belangrijk om te begrijpen hoe de hersenen werken, vooral in psychiatrische ziekten en cognitieve taken zoals leren en geheugen. Met optogenetica kunnen de enige cellen of de celbevolking tot gehele kringen door licht worden gemoduleerd. Gemeenschappelijke optogenetische instrumenten zoals Channelrhodopsin2 (ChR2) of Archaerhodopsine (Arch) zijn in staat om neuronen te activeren of te dempen, of het vrijkomen van zenders te verhogen of te remmen bij axonterminals die projecteren op verschillende hersengebieden1,,2,,3,4. Arch moet echter zorgvuldig worden gebruikt, omdat werd aangetoond dat de activering ervan bij presynaptische terminals de spontane zenderreleaseverhoogt 5. Arch is een naar buiten corrigerende protonpomp die de pH-waarde in de cel verandert. Dit alkalische milieu induceert calciuminstroom en verbetert de zender release5. Om specifiek intracellulaire signaleringstrajecten te moduleren, kunnen receptorchimera’s die bestaan uit een lichtactiverend optogenetisch gereedschap, zoals rhodopsine of kegelopsine, in combinatie met een adequate G-eiwit gekoppelde receptor, worden gemaakt6,7,8. De hoeveelheid en variatie van beschikbare optogenetische gereedschappen is de afgelopen tien jaar aanzienlijk toegenomen.
Het doel van optogenetica is om neuronale circuits te manipuleren tijdens gedrag. Optogenetica maakt bijvoorbeeld het meten van acute gedragsveranderingen mogelijk, zoals veranderingen in angstgedrag. Optogenetische hulpmiddelen worden geleverd in doelgebieden van de hersenen via virale vectoren. Met behulp van speciale promotors en versterkers, of het Cre-loxP-systeem, kan celtype specificiteit worden gewaarborgd voor de expressie van optogenetische instrumenten(figuur 1A). Er zijn verschillende genetisch gemodificeerde muislijnen die het enzym Cre-Recombinase alleen in specifieke celtypen uitdrukken. Bijvoorbeeld, Nex-Cre muizen uitdrukken de Cre-Recombinase in piramidale neuronen in de cortex en de hippocampus onder de controle van de Nex-promotor10. Dit enzym is in staat om DNA-sequenties om te keren, die worden geflankeerd door loxP kanten11. Bijgevolg kan de DNA-sequentie van een dubbel floxed optogenetic tool, die wordt omgekeerd en geflankeerd door loxP kanten, alleen worden getranscribeerd door neuronen die de Cre-Recombinasebezitten, maar niet door andere neuronale types12,13. In het geval van Nex-Cre muizen zal het optogenetische gereedschap uitsluitend worden uitgedrukt in piramidale neuronen. Lichtstimulatie van bepaalde hersengebieden wordt dan bereikt via chronische implantatie van optische vezels direct boven het gebied van belang. Dieren kunnen dan worden gekoppeld aan een geschikte lichtbron en zich vrij gedragen in bijna allerlei gedragstesten.
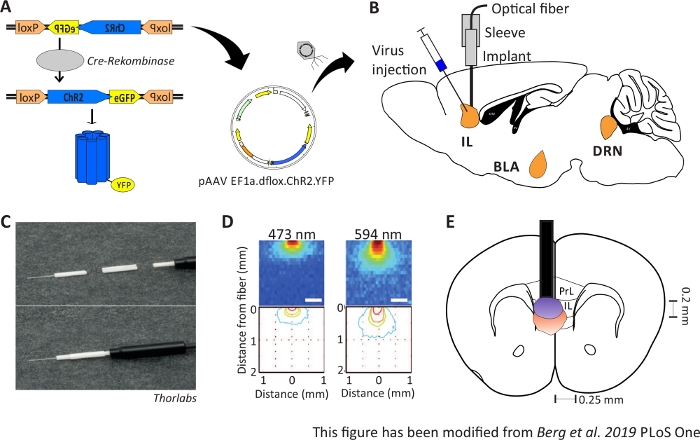
Figuur 1: Injectie en implantatie. A) Cre-loxP systeem voor ChR2-YFP. Dubbele floxed optogenetic tool is verpakt in een adeno geassocieerd virus (AAV) voor injectie in het hersenweefsel. B) Sagitttal view of the virus injection and implantation of an optical neuronal interface into/above the IL region of the mPFC. Injectie en implantatie werden gedaan van bovenaf. Alle regio’s van belang, IL, BLA en DRN, worden getoond. C) Gedetailleerde weergave van de geïmplanteerde glasvezel, mouw en lichtbron. D) Verspreiding van blauwe en rode laserlichtstimulatie in grijsstof hersenweefsel van een lichtvezel van 200 μm (Yizhar et al. 2011). Blauw licht verspreidt zich maximaal 0,5 mm in het weefsel, rood licht ongeveer 1 mm. Kleurcodering: rood 50%, geel 10%, groen 5%, blauw 1% als het licht dit gebied bereikt. E) Coronale weergave van de eenzijdige implantatie direct boven de linker IL met een 200 μm optische vezel. Het IL-gebied heeft een breedte van 0,25 mm per halfrond en een diepte van 0,2 mm. Blauwe en rode lampen zijn de boarder van 5% licht verspreiden en worden overgebracht van Yizhar et al. naar de juiste grootte. LoxP: locus van X-over P1; ChR2: Channelrhodopsine; YFP: geel fluorescerend eiwit; dflox: dubbel gevlobbel; IL: infralimbic cortex; BLA: basolaterale amygdala; DRN: dorsale raphe kernen; PrL: prelimbic regio. Dit cijfer is gewijzigd ten opzichte van Berg 201948. Klik hier om een grotere versie van dit cijfer te bekijken.
Optogenetische benaderingen worden gebruikt omdat het zowel hoge temporele als ruimtelijke resolutie14 en celtypespecifieke modulatie mogelijk maakt. Bovendien is het mogelijk om het geïmplanteerde apparaat herhaaldelijk te gebruiken zonder verdere behandeling. Na een stereotactische operatie, waarbij de injectie van een adeno-geassocieerd virus met het optogenetische gereedschap en de implantatie van de optische vezel wordt uitgevoerd, kunnen muizen twee weken herstellen. We hebben gekozen voor een hersteltijd van slechts 2 weken, omdat dit genoeg tijd is om te herstellen van de operatie en voor het virus om uit te drukken. Aangezien de gedragsexperimenten worden gevolgd door immunohistochemie, moeten we ervoor zorgen dat muizen niet te oud worden tijdens het experiment; anders wordt de weefselkwaliteit verlaagd. Ze tonen geen duidelijke gedragsstoornissen van het implantaat en nemen deel aan typisch kooigedrag. Natuurlijk gaat de implantatie gepaard met een aanzienlijke chirurgische laesie; daarom worden de muizen intensief gemonitord. Na de operatie moeten muizen alleenstaand zijn, omdat groepsmuizen elkaars verse wonden en implantaten hebben verwond. Echter, huisvestingsomstandigheden hebben een grote invloed op het angstniveau van mannelijke muizen, als single gehuisvest muizen tonen lagere angstniveaus15 en in het algemeen minder depressieve-achtige symptomen16.
Chemische of elektrische manipulatie van hersencircuits ontbreekt de hoge cel type specificiteit van optogenetica en hebben een lagere temporele en ruimtelijke resolutie14,17,18. Afhankelijk van de experimentele vraag kan elektrische of chemische stimulatie verschillende voordelen hebben. Bij het passeren van glasvezelterminals in een specifieke regio moet ook worden gestimuleerd, elektrische stimulatie is de beste methode. Chemische stimulatie is een goede keuze voor wanneer zenderspecifieke receptoren in een hele regio door agonisten moeten worden geactiveerd. Een ander groot voordeel van optogenetica in vergelijking met chemische of elektrische stimulatie is dat endogene, neuronen zijn niet gevoelig voor licht, die het optreden van bijwerkingenvermijdt 19. Inderdaad, hoge lichtintensiteiten kunnen leiden tot verwarmingseffecten8,20, maar als gevolg van de juiste controlegroepen kunnen de gedragseffecten als gevolg van optogenetische manipulatie worden geëlimineerd.
Het onderzoeken van knaagdiergedrag, vooral met betrekking tot psychiatrische ziekten, is sterk verbeterd met optogenetica bij vrij bewegende dieren, omdat het de directe modulatie van enkele receptoren tot specifieke celpopulaties21 en circuits22mogelijk maakt. De mogelijkheid om de acute effecten van dergelijke modulaties te meten, evenals de gedragseffecten op lange termijn na een bepaalde tijd23 of na chronische stimulatie24,maakt een brede flexibiliteit van experimentele ontwerpen mogelijk en biedt zeer gedetailleerde inzichten in hersencircuits. Lichtstimulatie kan worden gebruikt om neuronen op de injectieplaats van het optogenetische gereedschap te moduleren. Wanneer zowel de injectie als de implantatie hetzelfde hersengebied aanpakken, kunnen cellichamen en rugprojecterende axonen van principiële neuronen en interneuronen in deze regio worden gericht op3,6,8. Echter, de lichtvezel kan ook worden geïmplanteerd in een andere regio dan de geïnjecteerde. In dit geval kan lichtstimulatie het vrijkomen van zenders moduleren bij axonterminals in projectiegebieden van het geïnjecteerde gebied25,26,27.
In de studie hier wordt optogenetica gebruikt in combinatie met experimenten om angstgerelateerd gedrag te analyseren. Angstgerelateerde psychiatrische ziekten treffen meer dan een derde van de wereldbevolking28,29,30 en veroorzaken een hoge economische last31. De getroffenen hebben last van een gevoel van opwinding, spanning en zorgen, gevolgd door vermijdingsgedrag32,33. Deze chronisch voorkomende negatieve emoties, die vooral gericht zijn op toekomstige gebeurtenissen34, sterk interfereren met het dagelijks leven van patiënten. Gemeenschappelijke behandelingen zoals benzodiazepines of selectieve serotonine heropname remmers (SSRIs) zijn alleen succesvol bij sommige van de patiënten. Een groot aantal mensen reageert helemaal niet op de behandeling35, waaruit blijkt dat het mechanisme dat aan dergelijke ziekten ten grondslag ligt nog niet volledig is begrepen. Het is bekend dat de mediale prefrontale cortex (mPFC) een belangrijke rol speelt bij de ontwikkeling en manifestatie van angst21,25,27,36,37,38. Met name de overactivering van het infralimbic cortex (IL) gebied in het mPFC kan deel uitmaken van angstgerelateerde aandoeningen39,40. Het hier beschreven voorbeeldexperiment kan helpen om te begrijpen hoe modulaties in het IL-gebied van het mPFC angstgedrag beïnvloeden. Bovendien kan de ontwikkeling van nieuwe therapeutische strategieën voor angstgerelateerde psychiatrische ziekten mogelijk ook worden ondersteund.
2-6 maanden oude mannelijke Nex-Cre muizen worden gebruikt om ChR2 specifiek uit te drukken in piramidale neuronen binnen de IL regio van de mPFC41. Nex-Cre muizen hebben een C57Bl/6 achtergrond en drukken het enzym Cre-recombinase specifiek uit in piramidale neuronen. Tijdens een stereotactische operatie wordt dubbel floxed ChR2-DNA via adeno-geassocieerde virale vectoren in het IL-gebied geïnjecteerd. Het optische implantaat wordt direct boven het interessegebied(figuur 1B)geplaatst en het implantaat wordt gefixeerd met tandcement. Controledieren krijgen een injectie met dubbel floxed tdTomato-DNA in dezelfde regio om celspecifieke expressie na te bootsen.
Dieren zijn groepshuis tot de dag van de operatie en daarna zijn single gehuisvest om verwondingen van andere muizen te voorkomen. Muizen zijn ondergebracht in individuele geventileerde kooi (IVC) rekken in TypI-L kooien voor eenpersoonsmuizen. De licht-donkere cyclus volgt een 12:12 uur ritme, de lichtfase vanaf 10 uur. Alle gedragsexperimenten worden uitgevoerd in de donkere fase, die lijkt op de actieve fase van knaagdieren. Water en standaard voedsel pellets zijn beschikbaar ad libitum. Na twee weken herstel, wat zorgt voor een voldoende expressie van ChR2 in piramidale neuronen, worden muizen gebruikt voor gedragsexperimenten.
Het Open Field (OF) is een 50 cm x 50 cm kwadraat doolhof met gezandstraalde 40 cm hoge muren. De grond is verdeeld in 16 vierkanten waar de binnenste 4 het centrum vertegenwoordigen. Het gemeten gedrag is: 1) tijd doorgebracht in het centrum, 2) aantal middelste inzendingen, en 3) totale afstand verplaatst. Tijdens dit experiment zijn er 4 proeven in totaal 20 minuten. In proeven 1 en 3 treedt geen lichtstimulatie op en in proeven 2 en 4 wordt een 20 Hz-stimulatie met 5 ms lichtpuls en 1 mW lichtintensiteit van 473 nm uitgevoerd(figuur 2A). In de latere proeven werd rekening gehouden met gewenning aan het testgebied, maar het gebruik van met schijn geïnjecteerde bestrijdingsdieren geeft aan hoe gewenning wordt uitgedrukt.
De Barnes Maze is een experiment voor leren en geheugen. Het is een cirkelvormig platform met een diameter van 92 cm en bevat 20 gelijke gaten rond de omtrek. 19 van de gaten zijn gesloten en onder één gat wordt een vluchtdoos voorgesteld. Gedurende 4 opeenvolgende dagen hebben muizen 4 trainingsproeven om de locatie van de escape box te leren. Op de5e dag wordt de escape box verwijderd en worden muizen getest op hoeveel tijd ze nodig hebben om het juiste gat te vinden. Het gemeten gedrag is: 1) Tijd tot de vluchtdoos/correct gat wordt gevonden, 2) Aantal doelbezoeken en fouten, en 3) De afstand bewogen tot in het vluchtvak. De lichtstimulatie in verschillende groepen gebeurt tijdens de verwerving of consolidatie, die plaatsvinden op de trainingsdagen 1-4, of tijdens het ophalen op de testdag, die dag 5 is (Figuur 2D).
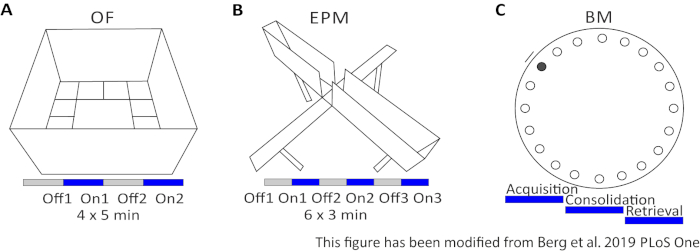
Figuur 2: Gedragsexperimenten met optogenetische protocollen. A) Schematische tekening van het Open Veld-experiment met het bijbehorende lichtstimulatieprotocol. C) Schematische tekening van het Elevated-Plus Doolhof experiment met het bijbehorende lichtstimulatieprotocol. D) Schematische tekening van de Barnes Maze experimenteren met de bijbehorende lichtstimulatie protocol. EPM: Elevated-Plus Doolhof; VAN: Open veld; BM: Barnes Maze Test. Dit cijfer is gewijzigd ten opzichte van Berg 201948. Klik hier om een grotere versie van dit cijfer te bekijken.
Voor optogenetische stimulatie moeten de lichtintensiteit en frequentie worden aangepast aan het optogenetische gereedschap en het neuronale type dat wordt onderzocht. De laagst mogelijke lichtintensiteit moet worden gebruikt om schade aan het weefsel te voorkomen, aangezien verschillende studies hebben aangetoond dat er mogelijke verwarmingseffecten zijn als gevolg van een sterke lichtintensiteit8,20. Voor ChR2 wordt een 20 Hz stimulatie met een 5 ms lichtpuls vaak gebruikt2. Omdat ChR2 vrij lichtgevoelig is, is 1 mW lichtintensiteit voldoende. Het lichtstimulatieprotocol wisselt af tussen licht uit en op proeven om gedragsveranderingen direct te meten. De externe ruimtevoorwaarden voor gedragsexperimenten moeten stabiel blijven voor de hele groep dieren. Belangrijke omstandigheden om te overwegen zijn het lawaai (houd er rekening mee dat apparaten zelf geluid kunnen maken), de geur (reinig altijd de gedragsopstellingen met ethanol), de lichtintensiteit en de onderzoeker. De experimentator moet altijd dezelfde persoon zijn. Bovendien moet het tijdstip van de experimenten hetzelfde zijn voor alle dieren in één groep, een paar uur na het begin van de donkere fase in de faciliteit heeft de voorkeur.
Het doel van dit experiment is het verhogen van de excitatie/remming (E/I) verhouding in het IL-gebied door sterke activering van excitatory piramidale neuronen. Een verbeterde E/I-verhouding in dit speciale cortexgebied is bekend dat het de angstniveaus bij muizenverhoogt 40,42,43,44.
