Merkezi sinir sisteminde nöronal devrelerin modülasyonu ve davranışsal sonuçları beynin nasıl çalıştığını anlamak için önemlidir, özellikle psikiyatrik hastalıklar ve öğrenme ve hafıza gibi bilişsel görevler. Optogenetik ile, tek hücre veya hücre popülasyonları kadar tüm devreler ışık tarafından modüle edilebilir. Channelrhodopsin2 (ChR2) veya Archaerhodopsin (Arch) gibi yaygın optogenetik araçlar nöronları etkinleştirmek veya susturmak, ya da artırmak veya farklı beyin bölgelerine projektör terminallerinde verici salınımı inhibe edebiliyoruz1,2,3,4. Ancak, Arch presinaptik terminallerde aktivasyonu spontan vericisalınımınıartırır gösterildiği gibi dikkatle kullanılması gerekir 5 . Arch, hücre içindeki pH değerini değiştiren dışa doğru düzeltici bir proton pompasıdır. Bu alkali ortam kalsiyum akını neden olur ve verici serbestartırır 5. Özellikle hücre içi sinyal yollarını modüle etmek için, rodopsin veya koni opsin gibi hafif bir aktive optogenetik araçtan oluşan reseptör kimeraları, yeterli bir G-protein birleştirilmiş reseptör ile birlikte, oluşturulabilir6,7,8. Mevcut optogenetik araçların miktarı ve varyasyonu son on yılda önemli ölçüde artmıştır9.
Optogenetik amacı davranış sırasında nöronal devreleri manipüle etmektir. Optogenetik, örneğin anksiyete davranışındaki değişiklikler gibi akut davranış değişikliklerinin ölçülmesini sağlar. Optogenetik aletler viral vektörler aracılığıyla beynin hedef bölgelerine ulaştırıldı. Özel organizatörler ve arttırıcılar veya Cre-loxP sistemi yardımıyla, optogenetik araçların ekspresyonu için hücre tipi özgüllüğü sağlanabilir (Şekil 1A). Sadece belirli hücre tiplerinde Cre-Rekombinaz enzimini ifade eden birkaç genetiği değiştirilmiş fare hattı vardır. Örneğin, Nex-Cre fareler Nex-promotor kontrolü altında korteks ve hipokampus piramidal nöronlarda Cre-Rekombimaz ifade10. Bu enzim, loxP kenarları11ile çevrili DNA dizilerini tersine çevirebilir. Sonuç olarak, ters ve loxP taraf tarafından çevrili bir çift floxed optogenetik aracın DNA dizisi, sadece Cre-Rekombinazsahip nöronlar tarafından transkripsiyonu olabilir , ama diğer nöronal türleri tarafından12,13. Nex-Cre fareler durumunda, optogenetik araç sadece piramidal nöronlar ifade edilecektir. Bazı beyin bölgelerinin ışık stimülasyonu daha sonra optik liflerin doğrudan ilgi alanının üzerinde kronik implantasyonu ile elde edilir. Hayvanlar daha sonra uygun bir ışık kaynağına birleştiğinde ve serbestçe davranış testleri hemen hemen her türlü davranış.
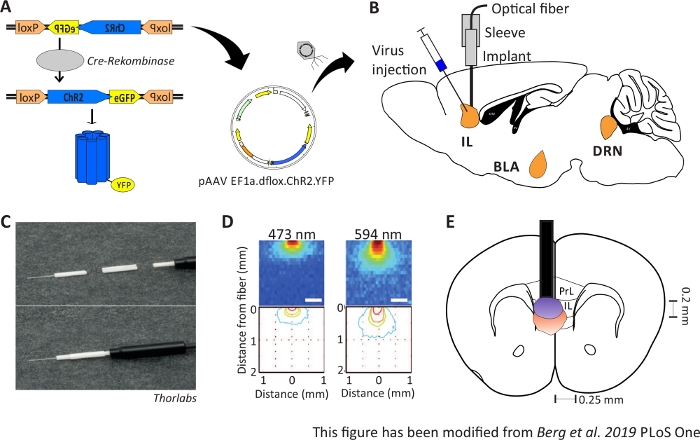
Şekil 1: Enjeksiyon ve implantasyon. A) ChR2-YFP için cre-loxP sistemi. Çift floxed optogenetik araç beyin dokusuiçine enjeksiyon için bir adeno ilişkili virüs (AAV) içinde paketlenir. B) MPFC’nin IL bölgesine/üstüne optik nöronal arayüz enjeksiyonunun ve implantasyonunun sagittal görünümü. Enjeksiyon ve implantasyon yukarıdan yapıldı. Tüm ilgi alanları IL, BLA ve DRN gösterilir. C)İmplante edilen optik fiber, kol ve ışık kaynağının ayrıntılı görünümü. D) Gri madde beyin dokusunda 200 μm ışık lifi ile mavi ve kırmızı lazer ışık stimülasyonunun yayılması (Yizhar ve ark. 2011). Mavi ışık, maksimum, doku içine 0,5 mm, yaklaşık 1 mm kırmızı ışık yayılır. Renk kodlaması: kırmızı %50, sarı %10, yeşil %5, ışık bu alana ulaşırsa mavi %1. E)200 μm optik fiber ile sol IL’nin hemen üzerinde tek taraflı implantasyonun koronal görünümü. IL bölgesi her yarımkürede 0,25 mm genişliğe ve 0,2 mm derinliğe sahiptir. Mavi ve kırmızı ampuller % 5 ışık yayılan yatılı ve Doğru boyuta Yizhar ve ark aktarılır. LoxP: X-over P1 locus; ChR2: Channelrhodopsin; YFP: sarı floresan protein; dflox: çift floxed; IL: infralimbic korteks; BLA: basolateral amigdala; DRN: dorsal raphe çekirdekleri; PrL: prelimbic bölge. Bu rakam Berg 201948’dendeğiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Hem yüksek zamansal hem de mekansal çözünürlük14 ve hücre tipine özgü modülasyon sağlayan optogenetik yaklaşımlardan yararlanılır. Ayrıca, implante edilen cihazı daha fazla tedavi edilmeden tekrar tekrar kullanmak mümkündür. Optogenetik alettaşıyan adeno-ilişkili bir virüsün enjeksiyonu ve optik fiber inplantasyonunun yapıldığı stereotaktik cerrahiden sonra fareler iki hafta boyunca iyileşebilir. Biz sadece 2 haftalık bir iyileşme süresi seçtik, bu ameliyat tan kurtarmak için yeterli zaman çünkü ve virüs ifade etmek için. Davranışsal deneyler immünohistokimya tarafından takip edilir gibi, fareler deney sırasında çok yaşlı olsun emin olmak gerekir; aksi takdirde doku kalitesi azalır. İmplanttan belirgin davranış bozuklukları göstermez ler ve tipik kafes davranışlarına girişinler. Tabii ki, implantasyon önemli bir cerrahi lezyon eşlik eder; bu nedenle, fareler yoğun bir şekilde izlenir. Ameliyattan sonra, grup fareler birbirlerinin taze yaraları ve implantları zarar eğilimindedir gibi, fareler, tek ev olması gerekir. Ancak, konut koşulları erkek farelerin anksiyete düzeyi üzerinde büyük bir etkisi var, tek ev fareler düşük anksiyete düzeyleri göstermek gibi15 ve genel olarak daha az depresif benzeri belirtiler16.
Beyin devrelerinin kimyasal veya elektriksel manipülasyon optogenetik yüksek hücre tipi özgüllüğü eksikliği ve daha düşük bir zamansal ve mekansal çözünürlükvar 14,17,18. Deneysel soruya bağlı olarak, elektrikveya kimyasal stimülasyon farklı avantajlara sahip olabilir. Belirli bir bölgede lif terminalleri geçerken de uyarılması gerekir, elektrikstimülasyon en iyi yöntemdir. Kimyasal stimülasyon, tüm bölgedeki vericiye özgü reseptörlerin agonistler tarafından aktive edilmesi için iyi bir seçimdir. Optogenetik kimyasal veya elektrikstimülasyon ile karşılaştırıldığında bir diğer büyük avantajı, endojen, nöronlar ışığa duyarlı değildir, hangi yan etkilerin oluşumunu önler19. Gerçekten de, yüksek ışık yoğunlukları ısıtma etkileri neden olabilir8,20, ama uygun kontrol grupları nedeniyle, optogenetik manipülasyon nedeniyle davranışsal etkileri ortadan kaldırılabilir.
Kemirgen davranışlarını araştırmak, özellikle psikiyatrik hastalıklar açısından, serbestçe hareket eden hayvanlarda optogenetik ile büyük ölçüde düzelmiştir, belirli hücre popülasyonlarına kadar tek reseptörlerin doğrudan modülasyonu sağlar gibi21 ve devreler22. Bu tür modülasyonların akut etkilerini ölçme imkanı, ayrıca tanımlanmış bir süre23 veya kronik stimülasyon24sonra uzun vadeli davranışsal etkileri , deneysel tasarımları geniş bir esneklik sağlar ve beyin devreleri içine çok ayrıntılı anlayışlar sağlar. Işık stimülasyonu optogenetik aracın enjeksiyon yerinde bulunan nöronları modüle etmek için kullanılabilir. Enjeksiyon ve implantasyon aynı beyin bölgesine hitap ettiğinde, hücre organları ve bu bölgedeki ilke nöronve internöronların aksonları projelendirme3,6,,8hedeflenebilir. Ancak, ışık lifi de enjekte edilenden farklı bir bölgeye yerleştirilmesi olabilir. Bu durumda, ışık stimülasyonu enjekte edilen bölgenin projeksiyon alanlarında akson terminallerinde verici salınımını modüle edebilir25,26,27.
Burada yapılan çalışmada, optogenetik anksiyete ile ilgili davranışları analiz etmek için deneyler ile birlikte kullanılır. Anksiyete ile ilgili psikiyatrik hastalıklar dünya nüfusunun üçte birinden fazlasını etkiler28,29,30 ve yüksek bir ekonomik yük neden31. Bu etkilenen uyarılma hissi muzdarip, gerginlik ve endişe kaçınma davranışı takip32,33. Bu kronik olarak meydana gelen olumsuz duygular, esas olarak gelecekteki olaylar a odaklanmıştır34, güçlü hastaların günlük yaşam ını müdahale. Benzodiazepinler veya selektif serotonin geri alım inhibitörleri gibi yaygın tedaviler (SSRI) sadece bazı hastalarda başarılı. İnsanların büyük bir kısmı tedaviye35yaşında yanıt vermez , bu tür hastalıkların altında yatan mekanizmanın henüz tam olarak anlaşılamadığını gösterir. Medial prefrontal korteks (mPFC) anksiyete gelişimi ve tezahürü önemli bir rol oynadığı bilinmektedir21,25,27,36,37,38. Özellikle, mPFC infralimbic korteks (IL) bölgenin aşırı aktivasyonu anksiyete ile ilgili bozuklukların bir parçası olabilir39,40. Burada açıklanan örnek deney, mPFC’nin IL bölgesindeki modülasyonların anksiyete davranışını nasıl etkilediğini anlamanızda yardımcı olabilir. Ayrıca, anksiyete ile ilgili psikiyatrik hastalıklar için yeni tedavi stratejilerinin geliştirilmesi de potansiyel olarak desteklenebilir.
2-6 aylık erkek Nex-Cre fareler mPFC41IL bölge içinde piramidal nöronlar özellikle ChR2 ifade etmek için kullanılır. Nex-Cre fareler c57Bl/6 arka plana sahiptir ve özellikle piramidal nöronlarda Cre-rekombinaz enzimini ifade eder. Stereotaktik cerrahi sırasında, çift floxed ChR2-DNA adeno ilişkili viral vektörler ile IL bölgeye enjekte edilir. Optik implant ilgi alanının(Şekil 1B)üzerine yerleştirilir ve implant diş çimentosu ile sabitlenir. Kontrol hayvanları hücreye özgü ifadeyi taklit etmek için aynı bölgede çift floxed tdTomato-DNA enjeksiyonu alırlar.
Hayvanlar ameliyat gününe kadar grup içinde dirve daha sonra diğer farelerin yaralanmalarını önlemek için tek kişilik tir. Fareler tek ev fareler için TypI-L kafeslerde bireysel havalandırmalı kafes (IVC) raflarda yer almaktadır. Açık-koyu çevrim 12:12 saat ritmi, 10 başlayan ışık fazı izler. Tüm davranışsal deneyler, kemirgenlerin aktif evresini andıran karanlık evrede gerçekleştirilir. Su ve standart gıda peletleri reklam libitum mevcuttur. Piramidal nöronlarda ChR2’nin yeterli bir ekspresyonunu sağlayan iki haftalık iyileşmeden sonra, fareler davranışsal deneyler için kullanılır.
Açık Alan (OF), 50 cm x 50 cm kare lik bir labirent ve 40 cm yüksekliğinde kum püskürtmeli bir labirenttir. Zemin iç 4 merkezi temsil 16 kareler ayrılmıştır. Ölçülen davranış: 1) merkezde harcanan zaman, 2) merkez girişsayısı ve 3) toplam mesafe taşındı. Bu deney sırasında, toplam 20 dakika süren 4 deneme vardır. 1 ve 3 numaralı çalışmalarda ışık stimülasyonu yapılmaz ve 2 ve 4 numaralı çalışmalarda 5 ms ışık darbesi ve 1 mW ışık yoğunluğu 473 nm olan 20 Hz stimülasyon gerçekleştirilir(Şekil 2A). Daha sonraki çalışmalarda, test alanına alışma dikkate alınmıştır, ancak sahte enjekte kontrol hayvanlarının kullanımı nasıl alışkanlık ifade edilir gösterir.
Barnes Labirent öğrenme ve bellek için bir deneydir. Çapı 92 cm olan ve çevresi etrafında 20 eşit uzaklıkta delik bulunan dairesel bir platformdur. Deliklerden 19’u kapatılır ve bir deliğin altında bir kaçış kutusu sunulur. 4 gün üst üste, fareler kaçış kutusunun yerini öğrenmek için 4 eğitim denemeleri var. 5.th günde, kaçış kutusu kaldırılır ve fareler doğru deliği bulmak için ne kadar zamana ihtiyaçları olduğu test edilir. Ölçülen davranış: 1) Kaçış kutusu/doğru delik bulunana kadar olan süre, 2) Hedef ziyaret ve hata sayısı ve 3) Mesafe kaçış kutusuna kadar taşınır. Farklı gruplardaki ışık stimülasyonu ya 1-4 eğitim günlerinde gerçekleşen satın alma veya konsolidasyon sırasında ya da 5. gün olan test gününde geri alma sırasında yapılır(Şekil 2D).
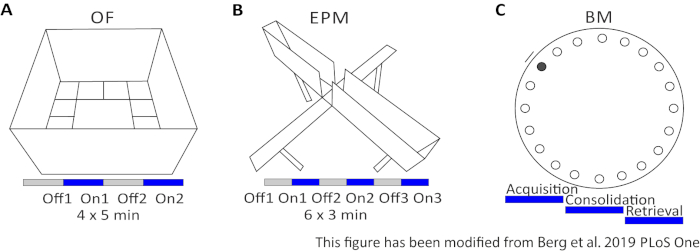
Şekil 2: Optogenetik protokollerle davranışsal deneyler. A) Açık Alan deneyinin ilgili ışık stimülasyon protokolü ile şematik çizimi. C) İlgili ışık stimülasyon protokolü ile Yükseltilmiş Artı Labirent deneyinşematik çizimi. D) Ilgili ışık stimülasyon protokolü ile Barnes Maze deney şematik çizim. EPM: Yükseltilmiş-Plus Labirent; OF: Açık Alan; BM: Barnes Labirent Testi. Bu rakam Berg 201948’dendeğiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Optogenetik stimülasyon için ışık yoğunluğu ve sıklığının araştırılan optogenetik araca ve nöronal tipe uyarlanmalıdır. Dokuya zarar vermemek için mümkün olan en düşük ışık yoğunluğu kullanılmalıdır, çünkü çeşitli çalışmalar güçlü ışık yoğunluğu nedeniyle olası ısıtma etkileri olduğunu göstermiştir8,20. ChR2 için, 5 ms ışık darbesi ile 20 Hz stimülasyon yaygın olarak kullanılır2. ChR2 ışığa oldukça duyarlı olduğundan, 1 mW ışık yoğunluğu yeterlidir. Işık stimülasyon protokolü, davranış değişikliklerini doğrudan ölçmek için ışık kapalı ve denemeler arasında değişir. Davranışsal deneyler için dış oda koşulları hayvanların tüm grup için istikrarlı kalmalıdır. Dikkate alınması gereken önemli koşullar gürültü (cihazların kendilerini gürültü yapabilir akılda tutmak), koku (her zaman etanol ile davranış kurulumları temiz), ışık yoğunluğu, ve deneyci. Deneyci her zaman aynı kişi olmalıdır. Ayrıca, deneylerin gün saati bir gruptaki tüm hayvanlar için aynı olmalıdır, tesiste karanlık fazBaşladıktan birkaç saat sonra tercih edilir.
Bu deneyin amacı, uyarıcı piramidal nöronların güçlü aktivasyonu ile IL bölgesindeki uyarma/inhibisyon (E/I) oranını artırmaktır. Bu özel korteks bölgesinde gelişmiş bir E / I oranı farelerde anksiyete düzeylerini artırmak için bilinmektedir40,42,43,44.
