La modulación de los circuitos neuronales en el sistema nervioso central y sus resultados conductuales son importantes para entender cómo funciona el cerebro, especialmente en enfermedades psiquiátricas y tareas cognitivas como el aprendizaje y la memoria. Con la optogenética, las células individuales o las poblaciones celulares hasta circuitos enteros pueden ser moduladas por la luz. Las herramientas optogenéticas comunes como Channelrhodopsin2 (ChR2) o Archaerhodopsin (Arch) son capaces de activar o silenciar las neuronas, o aumentar o inhibir la liberación del transmisor en terminales de axón que se proyectan a regiones cerebrales distintas1,,2,,3,,4. Sin embargo, Arch debe utilizarse cuidadosamente, ya que se demostró que su activación en terminales presinápticos aumenta la liberación espontánea del transmisor5. Arch es una bomba de protones rectificadora hacia afuera que cambia el valor de pH dentro de la célula. Este ambiente alcalino induce la afluencia de calcio y mejora la liberación del transmisor5. Para modular específicamente las vías de señalización intracelular, se pueden crear quimeras receptoras compuestas por una herramienta optogenética activable ligera, como rhodopsin o cono opsin, junto con un receptor acoplado adecuada a la proteína G,6,7,8. La cantidad y variación de las herramientas optogenéticas disponibles ha aumentado significativamente durante la última década9.
El propósito de la optogenética es manipular los circuitos neuronales durante el comportamiento. La optogenética permite, por ejemplo, la medición de cambios agudos de comportamiento, como cambios en el comportamiento de ansiedad. Las herramientas optogenéticas se entregan en las regiones diana del cerebro a través de vectores virales. Con la ayuda de promotores y potenciadores especiales, o el sistema Cre-loxP, se puede garantizar la especificidad del tipo de célula para la expresión de herramientas optogenéticas(Figura 1A). Hay varias líneas de ratón modificadas genéticamente que expresan la enzima Cre-Recombinase en tipos celulares específicos solamente. Por ejemplo, los ratones Nex-Cre expresan la Cre-Recombinase en neuronas piramidales en la corteza y el hipocampo bajo el control del Nex-promotor10. Esta enzima es capaz de invertir secuencias de ADN, que están flanqueadas por lados loxP11. En consecuencia, la secuencia de ADN de una herramienta optogenética de doble hilo uterino, que está invertida y flanqueada por lados loxP, sólo puede ser transcrita por neuronas que poseen la Cre-Recombinase,pero no por otros tipos neuronales12,,13. En el caso de los ratones Nex-Cre, la herramienta optogenética se expresará únicamente en neuronas piramidales. La estimulación lumítica de ciertas regiones cerebrales se logra a través de la implantación crónica de fibras ópticas directamente por encima de la región de interés. Los animales pueden entonces ser acoplados a una fuente de luz adecuada y comportarse libremente en casi todo tipo de pruebas de comportamiento.
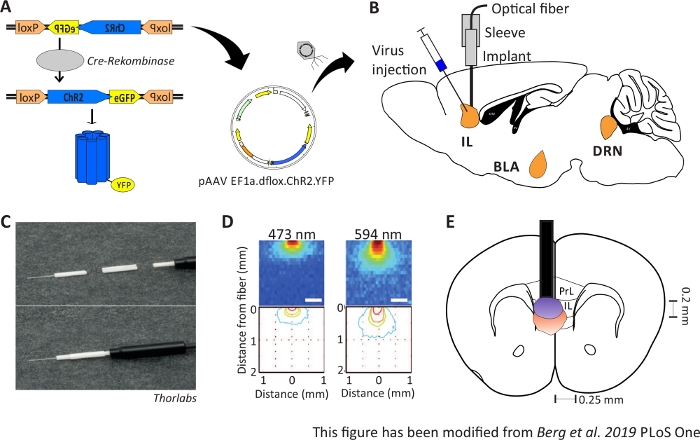
Figura 1: Inyección e implantación. A) Sistema Cre-loxP para ChR2-YFP. La herramienta optogenética de doble floxado se embala en un virus asociado al adeno (AAV) para inyección en el tejido cerebral. B) Vista sagitular de la inyección e implantación del virus de una interfaz neuronal óptica dentro/por encima de la región IL del mPFC. La inyección y la implantación se realizaron desde arriba. Se muestran todas las regiones de interés, IL, BLA y DRN. C) Vista detallada de la fibra óptica implantada, manguito y fuente de luz. D) Difusión de la estimulación de la luz láser azul y roja en el tejido cerebral de materia gris a partir de una fibra de luz de 200 m (Yizhar et al. 2011). La luz azul se extiende, como máximo, 0,5 mm en el tejido, luz roja de aproximadamente 1 mm. Codificación de color: rojo 50%, amarillo 10%, verde 5%, azul 1% si la luz llega a esta área. E) Vista coronal de la implantación unilateral directamente por encima de la IL izquierda con una fibra óptica de 200 m. La región IL tiene una anchura de 0,25 mm en cada hemisferio y una profundidad de 0,2 mm. Las bombillas azules y rojas son la placa de 5% de propagación de luz y se transfieren de Yizhar et al al al tamaño correcto. LoxP: locus de X-over P1; ChR2: Channelrhodopsin; YFP: proteína fluorescente amarilla; dflox: doble floxado; IL: corteza inmórbida; BLA: amígdala basolateral; DRN: núcleos de raphe dorsal; PrL: región prelímpico. Esta cifra ha sido modificada de Berg 201948. Haga clic aquí para ver una versión más grande de esta figura.
Se utilizan enfoques optogenéticos, ya que permite una alta resolución temporal y espacial14 y una modulación específica del tipo de celda. Además, es posible utilizar repetitivamente el dispositivo implantado sin tratamiento adicional. Después de una cirugía estereotáctica, donde se realiza la inyección de un virus adeno-asociado que lleva la herramienta optogenética y la implantación de la fibra óptica, los ratones pueden recuperarse durante dos semanas. Hemos elegido un tiempo de recuperación de sólo 2 semanas, porque este es el tiempo suficiente para recuperarse de la cirugía y para que el virus se exprese. Como los experimentos conductuales son seguidos por inmunohistoquímica, tenemos que asegurarnos de que los ratones no se envejezcas demasiado durante el experimento; de lo contrario, la calidad del tejido disminuye. No muestran impedimentos de comportamiento evidentes del implante y se involucran en el comportamiento típico de la jaula. Por supuesto, la implantación se acompaña de una lesión quirúrgica significativa; por lo tanto, los ratones son monitoreados intensivamente. Después de la cirugía, los ratones necesitan ser alojados en un solo lugar, ya que los ratones alojados en grupo tienden a herirse a las heridas e implantes frescos de los demás. Sin embargo, las condiciones de la vivienda tienen un gran impacto en el nivel de ansiedad de los ratones machos, ya que los ratones de una sola casa muestran niveles de ansiedad más bajos15 y en general síntomas menos depresivos16.
La manipulación química o eléctrica de los circuitos cerebrales carece de la alta especificidad del tipo celular de la optogenética y tiene una resolución temporal y espacial más baja14,17,18. Dependiendo de la pregunta experimental, la estimulación eléctrica o química puede tener diferentes ventajas. Cuando también es necesario estimular los terminales de fibra en una región específica, la estimulación eléctrica es el mejor método. La estimulación química es una buena opción para cuando los receptores específicos del transmisor en toda una región deben ser activados por agonistas. Otra gran ventaja de la optogenética en comparación con la estimulación química o eléctrica es que endógenamente, las neuronas no son sensibles a la luz, lo que evita la aparición de efectos secundarios19. De hecho, altas intensidades de luz pueden inducir efectos de calentamiento8,20, pero debido a los grupos de control adecuados, los efectos conductuales debido a la manipulación optogenética pueden ser eliminados.
La investigación del comportamiento de los roedores, especialmente en lo que respecta a las enfermedades psiquiátricas, ha mejorado considerablemente con la optogenética en animales que se mueven libremente, ya que permite la modulación directa de receptores individuales hasta poblaciones celulares específicas21 y circuitos22. La posibilidad de medir los efectos agudos de tales modulaciones, así como los efectos conductuales a largo plazo después de un tiempo definido23 o después de la estimulación crónica24, permite una amplia flexibilidad de los diseños experimentales y proporciona información muy detallada sobre los circuitos cerebrales. La estimulación de la luz se puede utilizar para modular las neuronas situadas en el lugar de inyección de la herramienta optogenética. Cuando tanto la inyección como la implantación abordan la misma región cerebral, los cuerpos celulares y los axones que se proyectan de principios de neuronas e interneuronas en esta región pueden dirigirsea 3,,6,,8. Sin embargo, la fibra ligera también se puede implantar en una región diferente de la inyectada. En este caso, la estimulación de la luz puede modular la liberación del transmisor en terminales de axón en áreas de proyección de la región inyectada25,26,27.
En el estudio aquí, optogenética se utiliza en combinación con experimentos para analizar el comportamiento relacionado con la ansiedad. Las enfermedades psiquiátricas relacionadas con la ansiedad afectan a más de un tercio de la población mundial28,29,30 y causan una alta carga económica31. Los afectados sufren de una sensación de excitación, tensión y preocupación seguida de comportamiento de evasión32,,33. Estas emociones negativas de origen crónico, que se centran principalmente en eventos futuros34,interfieren fuertemente con la vida diaria de los pacientes. Tratamientos comunes como benzodiazepinas o inhibidores selectivos de la recaptación de serotonina (ISRS) sólo son exitosos en algunos de los pacientes. Una gran cantidad de personas no responden al tratamiento en los35años, lo que demuestra que el mecanismo subyacente a tales enfermedades aún no se entiende completamente. La corteza prefrontal medial (mPFC) es conocida por desempeñar un papel importante en el desarrollo y manifestación de la ansiedad21,,25,27,36,37,38. Específicamente, la sobreactivación de la región de la corteza infralimbia (IL) en el mPFC podría ser parte de trastornos relacionados con la ansiedad39,,40. El experimento de ejemplo descrito aquí podría ayudar a entender cómo las modulaciones en la región IL del mPFC influyen en el comportamiento de ansiedad. Además, el desarrollo de nuevas estrategias terapéuticas para enfermedades psiquiátricas relacionadas con la ansiedad también puede ser apoyado potencialmente.
Los ratones Nex-Cre macho de 2-6 meses de edad se utilizan para expresar ChR2 específicamente en neuronas piramidales dentro de la región IL del mPFC41. Los ratones Nex-Cre tienen un fondo C57Bl/6 y expresan la enzima Cre-recombinase específicamente en las neuronas piramidales. Durante una cirugía estereotáctica, se inyecta ADN ChR2 de doble flox en la región de IL a través de vectores virales asociados con adeno. El implante óptico se coloca directamente encima de la región de interés (Figura 1B) y el implante se fija con cemento dental. Los animales de control reciben una inyección de tdTomato-DNA de doble flox en la misma región para imitar la expresión específica de la célula.
Los animales son alojados en grupo hasta el día de la cirugía y después son alojados en una sola casa para evitar lesiones de otros ratones. Los ratones se alojan en bastidores individuales de jaula ventilada (IVC) en jaulas TypI-L para ratones de una sola casa. El ciclo luz-oscuridad sigue un ritmo de 12:12 h, la fase de luz a partir de las 10 AM. Todos los experimentos conductuales se realizan en la fase oscura, que se asemeja a la fase activa de los roedores. El agua y los pellets de alimentos estándar están disponibles ad libitum. Después de dos semanas de recuperación, lo que garantiza una expresión suficiente de ChR2 en las neuronas piramidales, los ratones se utilizan para experimentos conductuales.
El Campo Abierto (OF) es un laberinto cuadrado de 50 cm x 50 cm con paredes de 40 cm de altura. El suelo se divide en 16 cuadrados donde el interior 4 representa el centro. El comportamiento medido es: 1) tiempo invertido en el centro, 2) número de entradas centrales y 3) distancia total movida. Durante este experimento, hay 4 ensayos que suman 20 minutos. En los ensayos 1 y 3, no se produce estimulación lumínica, y en los ensayos 2 y 4, se realiza una estimulación de 20 Hz con pulso de luz de 5 ms y 1 mW de intensidad de luz de 473 nm (Figura 2A). En los ensayos posteriores, se tuvo en cuenta la habituación en el área de prueba, pero el uso de animales de control inyectados falsas indican cómo se expresa la habituación.
El Laberinto de Barnes es un experimento para el aprendizaje y la memoria. Es una plataforma circular de 92 cm de diámetro y contiene 20 agujeros equidistantes alrededor de la circunferencia. 19 de los agujeros están cerrados y debajo de un agujero se presenta una caja de escape. Durante 4 días consecutivos, los ratones tienen 4 ensayos de entrenamiento para aprender la ubicación de la caja de escape. El día 5, se retira la cajade escape, y se prueba a los ratones en cuanto a cuánto tiempo necesitan encontrar el agujero correcto. El comportamiento medido es: 1) Tiempo hasta que se encuentre el cuadro de escape/agujero correcto, 2) Número de visitas y errores de destino, y 3) Distancia movida hasta que se encuentre en el cuadro de escape. La estimulación lumítica en diferentes grupos se realiza ya sea durante la adquisición o consolidación, que tienen lugar en los días de entrenamiento 1-4, o durante la recuperación el día de la prueba, que es el día 5 (Figura 2D).
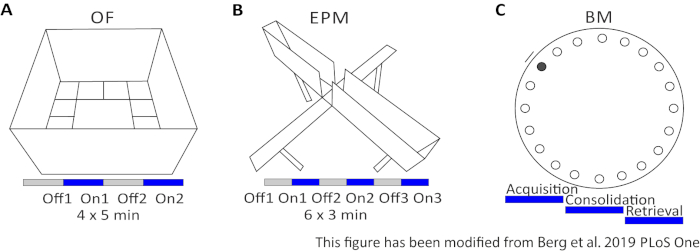
Figura 2: Experimentos conductuales con protocolos optogenéticos. A) Dibujo esquemático del experimento Campo Abierto con el protocolo de estimulación de luz correspondiente. C) Dibujo esquemático del experimento Elevated-Plus Maze con el protocolo de estimulación de la luz correspondiente. D) Dibujo esquemático del experimento Barnes Maze con el protocolo de estimulación de la luz correspondiente. EPM: Laberinto Elevado-Plus; OF: Campo abierto; BM: Prueba de laberinto de Barnes. Esta cifra ha sido modificada de Berg 201948. Haga clic aquí para ver una versión más grande de esta figura.
Para la estimulación optogenética, la intensidad y frecuencia de la luz deben adaptarse a la herramienta optogenética y al tipo neuronal que se está investigando. La menor intensidad lumínica posible debe utilizarse para evitar daños en el tejido, ya que varios estudios han demostrado que hay posibles efectos de calentamiento debido a la intensidad de la luz fuerte8,20. Para ChR2, una estimulación de 20 Hz con un pulso de luz de 5 ms se utiliza comúnmente2. Como ChR2 es bastante sensible a la luz, la intensidad de la luz de 1 mW es suficiente. El protocolo de estimulación de la luz alterna entre la luz apagada y los ensayos para medir directamente los cambios de comportamiento. Las condiciones externas de la habitación para los experimentos conductuales deben permanecer estables para todo el grupo de animales. Las condiciones importantes a tener en cuenta son el ruido (tenga en cuenta que los propios dispositivos pueden hacer ruido), el olor (limpiar siempre las configuraciones de comportamiento con etanol), la intensidad de la luz y el experimentador. El experimentador siempre debe ser la misma persona. Además, la hora del día de los experimentos debe ser la misma para todos los animales de un grupo, se prefiere unas horas después del inicio de la fase oscura en la instalación.
El objetivo de este experimento es aumentar la relación excitación/inhibición (E/I) en la región de IL a través de una fuerte activación de las neuronas piramidales excitatorias. Una relación E/I mejorada en esta región especial de la corteza se sabe para aumentar los niveles de ansiedad en ratones40,42,43,44.
