Moduleringen av neuronala kretsar i centrala nervsystemet och deras beteendemässiga resultat är viktiga för att förstå hur hjärnan fungerar, särskilt vid psykiatriska sjukdomar och kognitiva uppgifter som inlärning och minne. Med optogenetik kan enstaka celler eller cellpopulationer upp till hela kretsar moduleras av ljus. Vanliga optogenetiska verktyg som Channelrhodopsin2 (ChR2) eller Archaerhodopsin (Arch) kan aktivera eller tysta nervceller, eller öka eller hämma sändarutgåvan vid axonterminaler som projicerar till distinkta hjärnregioner1,2,3,4. Dock måste Arch användas försiktigt eftersom det visades att dess aktivering vid presynaptiska terminaler ökar spontan sändare release5. Arch är en utåt rättande protonpump som ändrar pH-värdet inuti cellen. Denna alkaliska miljö inducerar kalciumtillströmningen och förbättrar sändarutgåvan5. För att specifikt modulera intracellulära signalvägar, receptor chimeras består av ett ljus aktiveras optogenetiskt verktyg, såsom rhodopsin eller kon opsin, i samband med en adekvat G-protein kopplade receptor, kan skapas6,7,8. Mängden och variationen av optogenetiska verktyg som finns har ökat betydligt under det senaste decenniet9.
Syftet med optogenetik är att manipulera neuronal kretsar under beteende. Optogenetik möjliggör till exempel mätning av akuta beteendeförändringar såsom förändringar i ångestbeteende. Optogenetiska verktyg levereras till målregioner i hjärnan via virala vektorer. Med hjälp av särskilda tillskyndare och förstärkare, eller Cre-loxP-systemet, kan man säkerställa celltypsspecialitet för uttrycket av optogenetiska verktyg (Figur 1A). Det finns flera genetiskt modifierade muslinjer som uttrycker enzymet Cre-Recombinase i specifika celltyper endast. Till exempel, Nex-Cre möss uttrycka Cre-Rekombinas i pyramidala nervceller i cortex och hippocampus under kontroll av Nex-promotor10. Detta enzym är kompetent att invertera DNA-ordnar, som flankeras av loxP-sidor11. Följaktligen kan DNA-sekvensen av en dubbel-floxed optogenetiska verktyg, som är inverterad och flankerad av loxP sidor, endast transkriberas av nervceller som besitter Cre-Rekombinas, men inte av andra neuronala typer12,13. När det gäller Nex-Cre-möss kommer det optogenetiska verktyget enbart att uttryckas i pyramidala nervceller. Ljusstimulering av vissa hjärnregioner uppnås sedan via kronisk implantation av optiska fibrer direkt ovanför den region av intresse. Djur kan sedan kopplas till en lämplig ljuskälla och fritt bete sig i nästan alla typer av beteendemässiga tester.
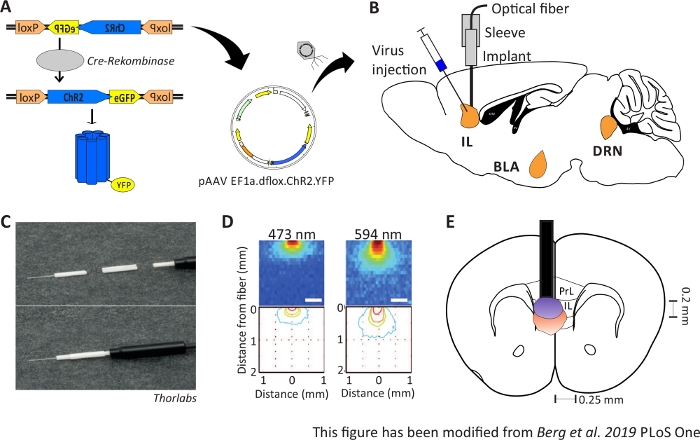
Bild 1: Injektion och implantation. A) Cre-loxP-system för ChR2-YFP. Dubbel floxed optogenetiska verktyget är förpackad i en adeno associerade virus (AAV) för injektion i hjärnvävnaden. B) Sagittal syn på virusinjektionen och implantationen av ett optiskt neuronalt gränssnitt in i/ovanför IL-regionen i mPFC. Injektion och implantation gjordes ovanifrån. Alla regioner av intresse, IL, BLA och DRN, visas. C) Detaljerad vy över den implanterade optiska fibern, hylsan och ljuskällan. D) Spridning av blå och röd laserljusstimulering i grå substans hjärnvävnad från en 200 μm ljusfiber (Yizhar et al. 2011). Blått ljus sprider sig, vid maximum, 0,5 mm in i vävnaden, rött ljus ca 1 mm. Färgkodning: röd 50%, gul 10%, grön 5%, blå 1% om ljus når detta område. E) Koronal syn på den ensidiga implantationen direkt ovanför vänster IL med en optisk fiber på 200 μm. IL-regionen har en bredd på 0,25 mm på varje halvklot och ett djup på 0,2 mm. Blå och röda glödlampor är boarder av 5% ljus spridning och överförs från Yizhar et al till rätt storlek. LoxP: locus av X-over P1; ChR2: Channelrhodopsin; YFP: gult fluorescerande protein; dflox: dubbel floxed; IL: infralimbic cortex; BLA: basolateral amygdala; DRN: dorsala raphe kärnor; PrL: prelimbic region. Denna siffra har ändrats från Berg 201948. Vänligen klicka här för att visa en större version av denna figur.
Optogenetiska tillvägagångssätt utnyttjas eftersom det möjliggör både hög tidsmässig och rumslig upplösning14 och celltyp specifik modulering. Dessutom är det möjligt att upprepat använda den implanterade enheten utan ytterligare behandling. Efter en stereotaktisk kirurgi, där injektionen av ett adeno-associerat virus som bär på det optogenetiska verktyget och implantationen av den optiska fibern utförs, kan möss återhämta sig i två veckor. Vi har valt en återhämtningstid på endast 2 veckor, eftersom detta är tillräckligt med tid att återhämta sig från operationen och för viruset att uttrycka. Eftersom beteendeexperimenten följs av immunohistokemi måste vi se till att möss inte blir för gamla under experimentet; annars minskas vävnadskvaliteten. De visar inga uppenbara beteendemässiga funktionsnedsättningar från implantatet och engagera sig i typiska bur beteende. Naturligtvis är implantationen åtföljd av en betydande kirurgisk lesion; därför övervakas mössen intensivt. Efter operationen måste möss enstaka inhysas, eftersom grupphusmöss tenderar att skada varandras färska sår och implantat. Men bostadsförhållanden har en stor inverkan på ångestnivån hos manliga möss, som enda inrymt möss visar lägre ångest nivåer15 och i allmänhet mindre depressiva-liknande symtom16.
Kemisk eller elektrisk manipulation av hjärnkretsar saknar optogenetikens höga celltypssärkalitet och har en lägre tidsmässig och rumslig upplösning14,17,18. Beroende på den experimentella frågan kan elektrisk eller kemisk stimulering ha olika fördelar. När passerar fiber terminaler i en specifik region måste också stimuleras, elektrisk stimulering är den bästa metoden. Kemisk stimulering är ett bra val för när sändarspecifika receptorer i en hel region ska aktiveras av agonister. En annan stor fördel med optogenetik jämfört med kemisk eller elektrisk stimulering är att endogent, nervceller är inte känsliga för ljus, som undviker förekomsten av biverkningar19. Faktum är att höga ljusintensiteter kan inducera värmeeffekter8,20, men på grund av rätt kontrollgrupper, de beteendemässiga effekter på grund av optogenetisk manipulation kan elimineras.
Undersökande gnagare beteende, särskilt i fråga om psykiatriska sjukdomar, har kraftigt förbättrats med optogenetik i fritt rörliga djur, eftersom det möjliggör direkt modulering av enstaka receptorer upp till specifika cellpopulationer21 och kretsar22. Möjligheten att mäta de akuta effekterna av sådana modulationer, liksom de långsiktiga beteendeeffekterna efter en definierad tid23 eller efter kroniskstimulering 24, möjliggör en bred flexibilitet av experimentella konstruktioner och ger mycket detaljerade insikter i hjärnkretsar. Ljusstimulering kan användas för att modulera nervceller som finns vid injektionsstället för det optogenetiska verktyget. När både injektion och implantation adress samma hjärnregionen, cell organ och tillbaka projicera axons av princip nervceller och interneurons i denna region kan riktas3,6,8. Men, den lätta fibern kan också implanteras i en region som skiljer sig från den injicerade. I detta fall kan ljusstimulering modulera sändarutgåva vid axonterminaler i projektionsområdena i den injicerade regionen25,26,27.
I studien här används optogenetik i kombination med experiment för att analysera ångestrelaterat beteende. Ångestrelaterade psykiatriska sjukdomar drabbar mer än en tredjedel av världens befolkning28,29,30 och orsakar en hög ekonomisk börda31. De som drabbas lider av en känsla av upphetsning, spänning och oro följt av undvikandebeteende 32,33. Dessa kroniskt förekommande negativa känslor, som huvudsakligen är inriktade på framtida händelser34, starkt störa det dagliga livet för patienter. Vanliga behandlingar som bensodiazepiner eller selektiva serotoninåterupptagshämmare (SSRI) är endast framgångsrika hos några av patienterna. En stor mängd människor svarar inte på behandlingen alls35, visar att den mekanism som ligger bakom sådana sjukdomar är ännu inte helt klarlagt. Den mediala prefrontala cortex (mPFC) är känd för att spela en viktig roll i utvecklingen och manifestation av ångest21,25,27,36,37,38. Specifikt kan överaktiveringen av den infralimbic cortex (IL) regionen i mPFC vara en del av ångest-relaterade sjukdomar39,40. Exemplet experimentet som beskrivs här skulle kunna bidra till att förstå hur modulationer i regionen IL i mPFC påverka ångest beteende. Dessutom kan utvecklingen av nya terapeutiska strategier för ångestrelaterade psykiatriska sjukdomar också potentiellt stödjas.
2-6 månader gamla manliga Nex-Cre möss används för att uttrycka ChR2 specifikt i pyramidala nervceller inom IL regionen av mPFC41. Nex-Cre möss har en C57Bl/6 bakgrund och uttrycka enzymet Cre-rekombinas specifikt i pyramidala nervceller. Under en stereotaktisk kirurgi injiceras dubbel floxed ChR2-DNA i IL-regionen via adeno associerade virala vektorer. Det optiska implantatet placeras direkt ovanför den region av intresse (Figur 1B) och implantatet är fast med tandcement. Kontroll djur får en injektion av dubbel floxed tdTomato-DNA i samma region att efterlikna cell specifika uttryck.
Djur är grupphus fram till operationsdagen och efteråt är enda inrymt för att undvika skador från andra möss. Möss är inhyst i enskilda ventilerade bur (IVC) rack i TypI-L burar för enkelhus möss. Den ljus-mörka cykeln följer en 12:12 h rytm, den ljusfas som börjar vid 10 AM. Alla beteendeexperiment utförs i den mörka fasen, som liknar den aktiva fasen av gnagare. Vatten och standardmatpellets finns ad libitum. Efter två veckors återhämtning, vilket säkerställer ett tillräckligt uttryck av ChR2 i pyramidala nervceller, används möss för beteendeexperiment.
Öppet fält (OF) är en 50 cm x 50 cm i kvadrat labyrint med sandblästrat 40 cm höga väggar. Marken är uppdelad i 16 rutor där den inre 4 representerar centrum. Det uppmätta beteendet är: 1) tid som tillbringas i centrum, 2) antal mittposter och 3) totalt avstånd flyttas. Under detta experiment, det finns 4 försök totalt 20 minuter. I försök 1 och 3 sker ingen ljusstimulering, och i försöken 2 och 4 utförs en 20 Hz-stimulering med 5 ms ljuspuls och 1 mW ljusintensitet på 473 nm (Figur 2A). I de senare försöken beaktades tillvänjning till testområdet, men användningen av skeninsprutade kontrolldjur anger hur tillvänjning uttrycks.
Barnes maze är ett experiment för inlärning och minne. Det är en cirkulär plattform som är 92 cm i diameter och innehåller 20 ekvidsliga hål runt omkretsen. 19 av hålen är stängda och under ett hål presenteras en utrymningsbox. För 4 dagar i följd har möss 4 träningsförsök för att lära sig platsen för utrymningslådan. På den5:e dagen tas utrymningsrutan bort, och möss testas på hur mycket tid de behöver för att hitta rätt hål. Det uppmätta beteendet är: 1) Tid tills utrymningsrutan/rätt hål hittas, 2) Antal målbesök och fel, och 3) Avstånd flyttat tills i utrymningsrutan. Ljusstimuleringen i olika grupper görs antingen under förvärv eller konsolidering, som sker på träningsdagarna 1-4, eller under hämtning på testdagen, som är dag 5 (Figur 2D).
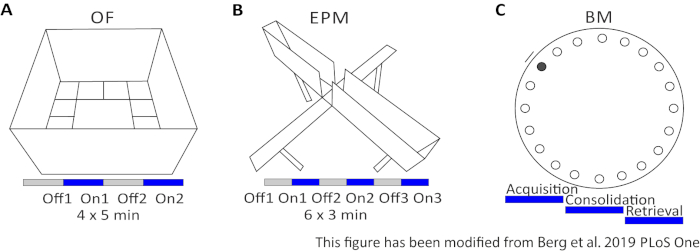
Figur 2: Beteendeexperiment med optogenetiska protokoll. A) Schematisk ritning av Open Field-experimentet med motsvarande ljusstimuleringsprotokoll. C) Schematisk ritning av Elevated-Plus Maze-experimentet med motsvarande ljusstimuleringsprotokoll. D) Schematisk ritning av Barnes Maze-experimentet med motsvarande ljusstimuleringsprotokoll. EPM: Förhöjd-Plus-labyrint; AV: Öppet sätta in; BM: Barnes labyrint test. Denna siffra har ändrats från Berg 201948. Vänligen klicka här för att visa en större version av denna figur.
För optogenetisk stimulering måste ljusintensiteten och frekvensen anpassas till det optogenetiska verktyg och neuronal typ som är under utredning. Lägsta möjliga ljusintensitet bör användas för att undvika skador på vävnaden, eftersom flera studier har visat att det finns möjliga uppvärmningseffekter på grund av stark ljusintensitet8,20. För ChR2 används vanligen en 20 Hz-stimulering med en 5 ms ljuspuls2. Som ChR2 är ganska ljuskänslig, är 1 mW ljusintensitet tillräcklig. Ljusstimuleringsprotokollet växlar mellan ljus från och på försök för att direkt mäta beteendeförändringar. De yttre rumsförhållandena för beteendeexperiment bör förbli stabila för hela gruppen av djur. Viktiga förutsättningar att tänka på är bullret (tänk på att enheterna själva kan göra ljud), lukten (rengör alltid beteendeuppställningarna med etanol), ljusintensiteten och experimentören. Försökspersonen ska alltid vara samma person. Dessutom bör tiden på dagen för experimenten vara densamma för alla djur i en grupp, några timmar efter starten av den mörka fasen i anläggningen är att föredra.
Målet med detta experiment är att öka excitation/ hämning (E / I) förhållandet i IL regionen genom stark aktivering av excitatoriska pyramidala nervceller. En förbättrad E / I-förhållande i denna speciella cortex regionen är känt för att öka ångest nivåer hos möss40,42,43,44.
