I fenotipi di crescita contribuiscono in modo critico alla forma fisica del lievito. La selezione naturale può distinguere in modo efficiente tra lignaggi con tassi di crescita diversi dall’inverso della dimensione effettiva della popolazione, che può superare i 108 individui1. Inoltre, la variabilità dei tassi di crescita tra gli individui all’interno di una popolazione è un parametro evolutivamente rilevante, in quanto può servire come base per strategie di sopravvivenza come la copertura dellepuntate 2,3,4,5,6. Pertanto, i test che consentono misurazioni altamente accurate dei fenotipi di crescita e delle loro distribuzioni sono fondamentali per lo studio dei microrganismi. Il saggio di crescita del microcolonio qui descritto può generare misurazioni individuali del tasso di crescita per ~10 5 microcolonie per esperimento. Questo saggio fornisce quindi un potente protocollo per studiare la genetica evolutiva e la genomica del lievito. Si presta particolarmente bene a testare come la variabilità all’interno delle popolazioni di singole cellule geneticamente identiche viene generata, mantenuta e contribuisce alla forma fisicadella popolazione 7,8,9,10.
Il metodo qui descritto (Figura 1) utilizza immagini a campo luminoso a basso ingrandimento catturate periodicamente di cellule che crescono in mezzi liquidi su una piastra inferiore in vetro da 96 o 384 pozze di vetro per tenere traccia della crescita in microcolonie. Le cellule aderiscono alla lettalina concanavalina A, che rivestise il fondo della piastra del microscopio e forma colonie bidimensionali. Poiché le microcolonie crescono in un monostrato, l’area della microcolonia è altamente correlata con la cella numero7. Pertanto, è possibile generare stime accurate del tasso di crescita del microcolony e del tempo di ritardo con un software di analisi delle immagini personalizzato che tiene traccia del tasso di cambiamento dell’area di ogni microcolony. Inoltre, la configurazione sperimentale può monitorare l’abbondanza e persino le localizzazioni subcellulari di proteine etichettate fluorescentmente espresse in queste microcolonie. L’elaborazione a valle dei dati di questo test di crescita della microcolonia può essere ottenuta mediante analisi personalizzate o con il software di analisi delle immagini esistente, come Processing Images Easily (PIE)11, un algoritmo per il riconoscimento robusto dell’area della colonia e l’analisi della crescita ad alta produttività da immagini a basso ingrandimento e luminose, disponibile tramite GitHub12.
Poiché le stime del tasso di crescita derivate dal saggio di crescita della microcolonia sono generate da un gran numero di misurazioni a colonia singola, sono estremamente accurate, con errori standard di diversi ordini di grandezza più piccoli delle stime stesse per un esperimento di dimensioni ragionevoli. Pertanto, il potere del saggio di rilevare differenze di tasso di crescita tra diversi genotipi, trattamenti o condizioni ambientali è elevato. Il formato multiwell-plate consente di confrontare numerose combinazioni di ambiente e genotipo diverse in un unico esperimento. Se i ceppi esprimono in modo costitutivo marcatori fluorescenti diversi, possono essere mescolati nello stesso pozzo e distinti dalla successiva analisi dell’immagine, che potrebbe aumentare ulteriormente la potenza consentendo la normalizzazione dei dati pozzo per pozzo.
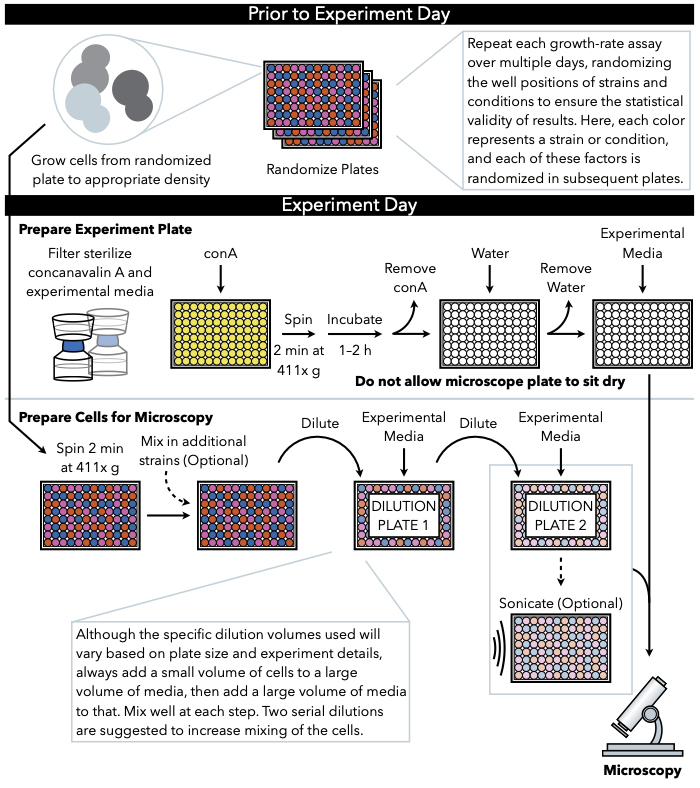
Figura 1: Rappresentazione schematica del protocollo. Questo protocollo segue due fasi principali, che sono la preparazione della piastra sperimentale e la preparazione delle cellule all’immagine. La randomizzazione delle piastre e la crescita delle cellule devono essere condotte prima e prima del giorno dell’esperimento. La miscelazione ripetuta delle cellule in ogni fase durante la diluizione è imperativa nei passaggi fino alla placcatura, e quindi si consiglia di preparare prima la piastra sperimentale in modo che sia pronta per la placcatura immediatamente dopo il completamento della diluizione cellulare. Clicca qui per visualizzare una versione più grande di questa figura.
