מחשוב דנ”א משתמש בערכה של אוליגונוקלאוטידים מעוצבים כמדיום לחישוב. אוליגונוקלאוטידים אלה מתוכנתים עם רצפים להרכבה דינמית על פי לוגיקה שצוינה על-ידי המשתמש ולהגיב לכניסות ספציפיות של חומצת גרעין. במחקרי הוכחת הרעיון, הפלט של החישוב מורכב בדרך כלל מסט של אוליגונוקלאוטידים שכותרתם פלואורסצנטית שניתן לזהות באמצעות אלקטרופורזה ג’ל או קוראי לוחות פלואורסצנטיות. במהלך 30 השנים האחרונות, מעגלים חישוביים DNA מורכבים יותר ויותר הודגמו, כגון מפלי לוגיקה דיגיטלית שונים, רשתותתגובהכימית, ורשתות עצביות 1,2,3. כדי לסייע בהכנת מעגלי DNA אלה, מודלים מתמטיים שימשו לחיזוי הפונקציונליות של מעגלי גנים סינתטיים4,5, וכלים חישוביים פותחו עבור עיצוב רצף DNA אורתוגונל6,7,8,9,10 . בהשוואה למחשבים מבוססי סיליקון, היתרונות של מחשבי DNA כוללים את יכולתם להתממשק ישירות עם ביומולקולים, לפעול בפתרון בהיעדר ספק כוח, כמו גם את הקומפקטיות והיציבות הכוללות שלהם. עם הופעתו של רצף הדור הבא, העלות של סינתזה של מחשבי DNA כבר יורדת בשני העשורים האחרונים בקצב מהר יותר מאשר חוק מור11. יישומים של מחשבים מבוססי DNA כאלה מתחילים כעת לצוץ, כגון לאבחון מחלות12,13, להפעלת ביופיסיקה מולקולרית14, וכפלטפורמות אחסון נתונים15.
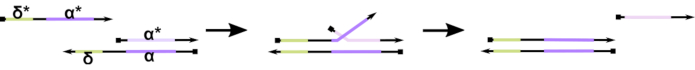
איור 1:מנגנון של תזוזת גדיל DNA בתיווך דריסת רגל. דריסת הבוהן, δ, היא רצף חופשי ולא מאוגד על דופלקס חלקי. כאשר תחום משלים (δ*) מוצג על גדיל שני, הדומיין δ החופשי משמש דריסת רגל להכלאה, ומאפשר לשאר הגדיל (ɑ*) לעקור לאט את המתחרה שלו באמצעות תגובה הפיכה רוכסת / בלתי מתפשרת המכונה העברת גדילים. ככל שאורך δ גדל, ה- ΔG לתגובה הקדמית פוחת, והעקירה מתרחשת ביתר קלות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
עד כה, רוב מחשבי הדנ”א משתמשים במוטיב מבוסס היטב בתחום הננוטכנולוגיה הדינמית של הדנ”א המכונה עקירת גדיל DNA בתיווך בוהן (TMDSD, איור 1)16. מוטיב זה מורכב דופלקס DNA כפול תקוע חלקית (dsDNA) המציגים overhangs קצרים “דריסת רגל” (כלומר, 7- עד 10 נוקלאוטידים (nt)). גדילי “קלט” חומצת גרעין יכולים לקיים אינטראקציה עם הדופלקסים החלקיים דרך דריסת המכהן. זה מוביל לעקירתו של אחד הגדילים מהדופלקס החלקי, והגדיל המשוחרר הזה יכול לשמש כקלט לדופלקסים חלקיים במורד הזרם. לכן, TMDSD מאפשרת סילוק את אותות ועיבוד מידע. באופן עקרוני, מוטיבים אורתוגונליים של TMDSD יכולים לפעול באופן עצמאי בפתרון, ומאפשרים עיבוד מידע מקביל. היו מספר וריאציות על תגובת TMDSD, כגון החלפת גדיל DNA בתיווך דריסת רגל (TMDSE)17, דריסות “ללא דליפות” עם תחומים ארוכים כפולים18, דריסות רצף לא תואמות19, ו “אחיזת יד”, עקירת גדילים בתיווך20. עקרונות עיצוב חדשניים אלה מאפשרים אנרגטיות ודינמיקה של TMDSD מכוונות יותר לשיפור ביצועי מחשוב ה- DNA.
מעגלי גנים סינתטיים, כגון מעגלי גנים תמלול, מסוגלים גם לחשב21,22,23. מעגלים אלה מוסדרים על ידי גורמי שעתוק חלבונים, המפעילים או מדחיקים שעתוק של גן על ידי קשירה לאלמנטים ספציפיים של DNA רגולטורי. בהשוואה למעגלים מבוססי דנ”א, למעגלי התמלול יש מספר יתרונות. ראשית, לתעתיק אנזימטי יש שיעור תחלופה גבוה בהרבה ממעגלי ה- DNA הקטליטיים הקיימים, ובכך יוצר יותר עותקים של פלט לכל עותק יחיד של קלט ומספק אמצעי יעיל יותר להגברת אותות. בנוסף, מעגלי שעתוק יכולים לייצר מולקולות פונקציונליות שונות, כגון aptamers או קידוד RNA שליח (mRNA) עבור חלבונים טיפוליים, כמו יציאות חישוב, אשר ניתן לנצל עבור יישומים שונים. עם זאת, מגבלה מרכזית של מעגלי התמלול הנוכחיים היא חוסר המדרגיות שלהם. הסיבה לכך היא שיש קבוצה מוגבלת מאוד של גורמי שעתוק מבוססי חלבון אורתוגונל, ועיצוב דה נובו של גורמי שעתוק חלבונים חדשים נשאר מאתגר מבחינה טכנית וגוזל זמן רב.
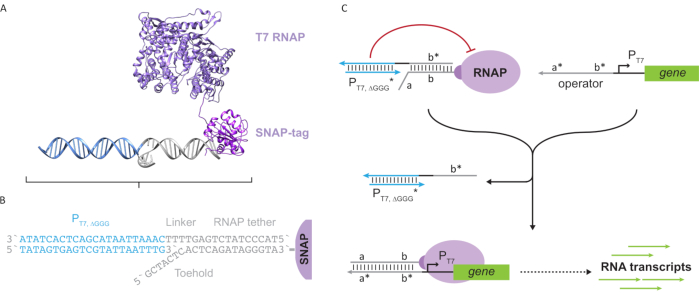
איור 2: הפשטה ומנגנון של קומפלקס פולימראז “לקשור” ו”כלוב”. (A ו- B) קשירה אוליגונוקלאוטיד מסומנת באופן אנזימטי לפולימראז T7 באמצעות תגובת SNAP-tag. כלוב המורכב ממקדם T7 “מזויף” עם תוחם משלים קשירה מאפשר לו להתיידד לקשירה ולחסום פעילות שעתוק. (C)כאשר המפעיל (a*b*) קיים, הוא נקשר לאיכות הבוהן על הרצועה אוליגונוקלאוטיד (ab) ומזיז את אזור b * של הכלוב, ומאפשר שעתוק להתרחש. נתון זה שונה מצ’ו ושי27. קיצורים: RNAP = RNA פולימראז. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
מאמר זה מציג אבן בניין חדשנית למחשוב מולקולרי המשלבת את הפונקציות של מעגלי שעתוק עם מדרגיות של מעגלים מבוססי DNA. אבן בניין זו היא T7 RNAP המחוברת באופן קוולנטי עם רצועת DNA חד-גדילית(איור 2A). כדי לסנתז את ה-T7 RNAP הזה, הפולימראז הותך לתגSNAP-24 של N-terminal והתבטא מחדש באשריצ’יה קולי. תג SNAP הגיב אז עם אוליגונוקלאוטיד פונקציונלי עם מצע BG. הרצועה אוליגונוקלאוטיד מאפשרת מיקום של אורחים מולקולריים בסמיכות לפולימראז באמצעות הכלאת DNA. אורח אחד כזה היה חוסם תמלול תחרותי המכונה “כלוב”, המורכב ודופלקס DNA “מזויף” של מקדם T7 ללא גנים במורד הזרם(איור 2B). כאשר הוא מאוגד לרנ”פ באמצעות קשירת האוליגונוקלאוטיד שלו, הכלוב מעכב פעילות פולימראז על ידי אי-התאמה של תבניות DNA אחרות עבור כריכת RNAP, ועיבוד ה-RNAP במצב “כבוי”(איור 2C).
כדי להפעיל את הפולימראז למצב “ON”, תוכננו תבניות DNA T7 עם תחומי “אופרטור” חד-גדיליים במעלה הזרם של מקדם T7 של הגן. תחום המפעיל (כלומר, תחום a*b* איור 2C) יכול להיות מתוכנן לעקור את הכלוב מה- RNAP באמצעות TMDSD ולמקם את ה- RNAP הקרוב למקדם T7 של הגן, ובכך ליזום שעתוק. לחלופין, תבניות DNA תוכננו גם כאשר רצף המפעיל היה משלים לגדילי חומצת גרעין עזר המכונים “גורמי שעתוק מלאכותיים” (כלומר, גדילי TFA ו- TFB באיור 3A). כאשר שני גדילים מוצגים לתוך התגובה, הם יתאספו באתר המפעיל, יצירת תחום פסאודו-רציף חדש a *b*. תחום זה יכול לאחר מכן לעקור את הכלוב באמצעות TMDSD כדי ליזום תמלול (איור 3B). גדילים אלה יכולים להיות מסופקים או אקסוגני או מיוצר.
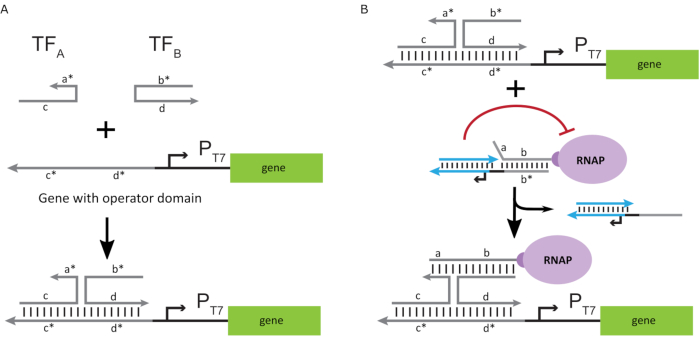
איור 3: תכנות סלקטיבי של פעילות פולימראז באמצעות מפעיל מתג בעל שלושה רכיבים. (A)כאשר קיימים גורמי התמלול (TFA ו- TFB), הם נקשרים לתחום המפעיל במעלה הזרם של המקדם, ויוצרים רצף פסאודו חד-גדילי (a*b*) המסוגל לעקור את הכלוב באמצעות תזוזת דנ”א מתווכת. (B)תחום a*b* זה יכול לעקור את הכלוב באמצעות TMDSD כדי ליזום תמלול. נתון זה שונה מצ’ו ושי27. קיצורים: TF = גורם שעתוק; RNAP = RNA פולימראז; TMDSD = תזוזת גדיל DNA בתיווך דריסת רגל. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
השימוש בגורמי שעתוק מבוססי חומצת גרעין עבור ויסות תמלול במבחנה מאפשר יישום מדרגי של התנהגויות מעגל מתוחכמות כגון לוגיקה דיגיטלית, משוב, ומדדי אותות. לדוגמה, ניתן לבנות מפלי שער לוגיים על ידי תכנון רצפי חומצות גרעין כך שהתעתיקים מגן במעלה הזרם מפעילים גן במורד הזרם. יישום אחד המנצל את המדורגים והמכפילים המסוגלים בטכנולוגיה מוצעת זו הוא פיתוח מעגלי מחשוב מולקולריים מתוחכמים יותר לאבחון נייד ועיבוד נתונים מולקולריים. בנוסף, שילוב יכולות המחשוב המולקולרי וסינתזת דה נובו RNA יכול לאפשר יישומים חדשים. לדוגמה, ניתן לתכנן מעגל מולקולרי כדי לזהות אחד או שילוב של RNAs המוגדרים על-ידי המשתמש כתשומות ופלט של RNAs טיפוליים או mRNAs המקודדים פפטידים פונקציונליים או חלבונים עבור יישומים רפואיים נקודתיים.
