تم تطوير تقنية التنميط الريبوسوم (RIBO-seq) في مختبر جوناثان ويسمان في جامعة كاليفورنيا ، سان فرانسيسكو1. بالمقارنة مع الأساليب الأخرى المستخدمة لدراسة التعبير الجيني على المستوى الترجمي ، يركز RIBO-seq على كل ربط ريبوسوم ل mRNA ويوفر معلومات حول موقعه والعدد النسبي للريبوسومات على نسخة. أنها تمكن من رصد عملية تخليق البروتين في الجسم الحي، ويمكن أن توفر دقة كودون واحد ودقة السماح لقياس كثافة الريبوسوم على حد سواء، مرنا الفردية وعلى طول النسخ الكامل في الخلية. في الأساس لتقنية ريبو-seq يكمن حقيقة أنه أثناء الترجمة يربط ريبوسوم جزيء مرنا وبالتالي يحمي جزء مدفون من النص من عملية الهضم ريبونوكليز. عند إضافة ريبونوكليز، يتم هضم الحمض النووي الريبي غير المحمي وتظل الشظايا المغلقة بالريبوسومات – التي يبلغ طولها عادة ~ 28-30 nt – سليمة. ويمكن بعد ذلك عزل هذه الشظايا، التي تسمى آثار أقدام ريبوسومية (RF)، وتسلسلها ورسم خرائط لها على النص الذي نشأت عنه مما أدى إلى الكشف عن الموضع الدقيق للريبوسومات. في الواقع، وقد استخدمت القدرة على حماية ريبوسوم شظايا مرنا منذ 1960s لدراسة الربط ribosomal ومواقع بدء الترجمة (TIS)2،3،4. ومع ذلك ، مع التقدم في تكنولوجيا التسلسل العميق ، أصبح RIBO-seq معيارا ذهبيا لرصد الترجمة5 ، والذي ، من خلال المشاركة الريبوسومية ، يمكن أن يوفر معلومات على نطاق الجينوم حول تخليق البروتين6. ريبوسوم التنميط سد الفجوة التكنولوجية التي كانت موجودة بين تحديد النسخ والبروتيوم6.
لإجراء التنميط ريبوسوم نحن بحاجة للحصول على lysate الخلية من الكائن الحي الذي نما في ظل الظروف التي تم التحقيق فيها. قد يؤدي تعطيل هذه الظروف أثناء جمع الخلايا وتحللها إلى توفير بيانات غير موثوقة. لمنع ذلك ، يتم استخدام مثبطات الترجمة والحصاد السريع وتجميد الفلاش في النيتروجين السائل. يمكن أن يتم التحلل الخلايا بواسطة طحن المبردة في التجانس الميكانيكية مثل مطحنة خلاط7،8 أو خافق حبة9، والتحوير من خلال ماصة10 أو مع إبرة11. يمكن إضافة المخزن المؤقت للتحلل قبل أو بعد وقت قصير من سحق الخلايا. في بروتوكولنا نستخدم النيتروجين السائل لقذائف الهاون والحشرات ، وكذلك أكسيد الألومنيوم كنهج ألطف لتعطيل جدار الخلية البكتيرية ، والذي يمنع قص الحمض النووي الريبي الذي غالبا ما يواجه عندما يتم تطبيق طرق مثل السونيشن. بعد السحق ، نضيف عازل تحلل بارد الجليد إلى المحتويات المبردة من قذائف الهاون. اختيار العازلة تحلل المناسبة مهم للحصول على أفضل قرار من آثار أقدام ريبوسومات. منذ قوة الأيونية يؤثر على كل من حجم RF ودقة إطار القراءة، فمن المستحسن حاليا لاستخدام المخازن المؤقتة تحلل مع قوة أيونية منخفضة والقدرة العازلة، حتى لو كان يبدو أن تكوين العازلة لا يؤثر على إشغال ريبوسومات على mRNAs11،12. المكونات الهامة للحاجز التحلل هي أيونات المغنيسيوم ، ووجود الذي يمنع تفكك الوحدات الفرعية ريبوسوم ويمنع التغيرات تشكيلية عفوية في ريبوسوماتالبكتيرية 11،13. أيونات الكالسيوم تلعب أيضا دورا هاما وضرورية لنشاط النيوكليز ميكروكوكال (MNase) المستخدمة في التنميط الريبوسوم البكتيرية طريقة14. إضافة جوانوسين 5′-[β,γ-imido]ثلاثي الفوسفات (GMP-PNP), التناظرية غير قابلة للتحلل من GTP, جنبا إلى جنب مع الكلورامفينيكول يمنع الترجمة خلال تحلل15.
عندما يتم الحصول على lysate، يتم توضيحه عن طريق الطرد المركزي وتقسيمها إلى قسمين، كل لريبو-seq وعالية الإنتاجية مجموع تسلسل مرنا (RNA-seq) لأنها تتم في وقت واحد(الشكل 1). يوفر RNA-seq نقطة مرجعية تمكن من مقارنة البيانات من كل من RIBO-seq و RNA-seq أثناء تحليل البيانات. يتم تعريف المترجم التحقيق من خلال تطبيع آثار أقدام ريبوسومات إلى وفرة مرنا16. يمكن أن تساعد البيانات من RNA-seq أيضا في تحديد الاستنساخ أو تسلسل القطع الأثرية17.
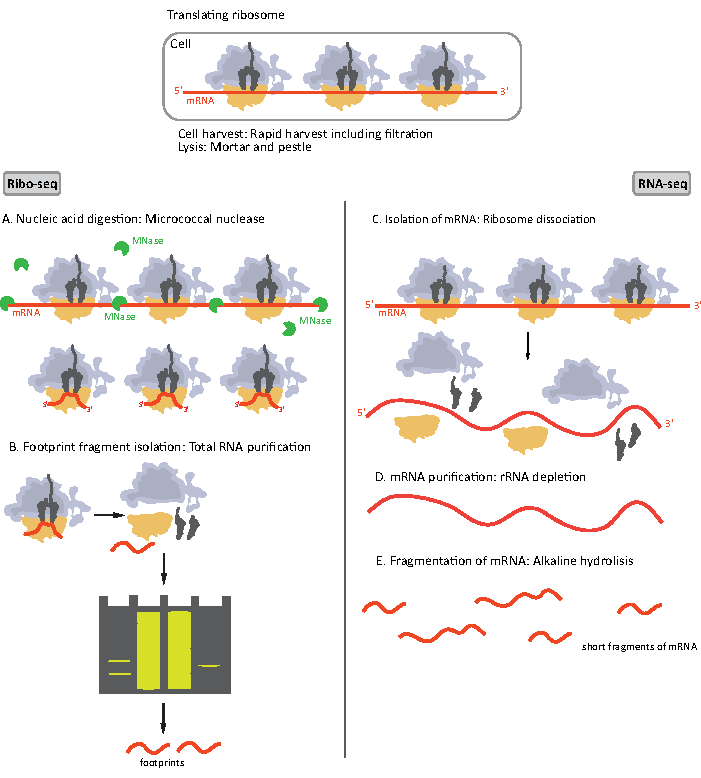
الشكل 1. تخطيطات إعداد عينة مرنا لريبو-seq و RNA-seq. لإعداد مكتبة ريبو-seq، يتم هضم الحمض النووي الريبي مع MNase (A)، تليها اختيار حجم RF من ~ 28-30 NT طول (B)؛ لرنا-seq يتم عزل (C)، استنفدت من rRNA (D)، ويتم تجزئة الحمض النووي الريبي الناتج عشوائيا إلى شظايا من أطوال متفاوتة (E). يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
الخطوات الأولية لإجراء إعداد عينة لريبو-seq و RNA-seq تختلف قليلا (الشكل 1). بالنسبة للتنميط الريبوسومي ، يحتاج الليسات إلى هضمه بواسطة إندونوكليز محدد لإضعاف جزيئات الحمض النووي الريبي غير المحمية بالريبوسومات. في البروتوكولات القياسية، يتم استرداد أحادية الحصول عليها عن طريق جهاز الطرد الفائق وسادة السكروز أو الطارد الفائق الانحدار السكروز8،14. في هذه المقالة، ونحن نظهر أن هذه الخطوة ليست ضرورية لعزل RF المطلوبة لريبو-seq في البكتيريا، وبالمثل للخلايا eukaryotic18،وأن اختيار حجم شظايا مرنا طول المناسبة من هلام البولي أكريلاميد كافية.
بالنسبة ل RNA-seq ، يتم الحصول على مرنا عن طريق استنفاد الحمض النووي الريبي من إجمالي الحمض النووي الريبي – جزيئات rRNA تهجين إلى مسابير أوليغونوكليوتيد البيوتينية التي ترتبط بالخرز المغناطيسي المغلف بالستريبتافيدين. ثم تتم إزالة مجمعات rRNA-oligonucleotide-حبة من العينة مع مغناطيس مما أدى إلى rRNA المنضب عينة19،20. ثم يتم تجزئة جزيئات الحمض النووي الريبي المنقى عشوائيا بواسطة التحلل المائي القلوية. يتم تحويل الأجزاء التي تم الحصول عليها من مرنا وكذلك آثار أقدام ريبوسومات إلى مكتبات cDNA وإعدادها لتسلسل عميق(الشكل 2). وهذا ينطوي على إصلاح ينتهي اللازمة بعد التحلل المائي القلوية (لرنا) والهضم الأنزيمي (لRF): dephosphorylation من 3 ‘ينتهي تليها الفوسفور من نهايات 5’. الخطوات التالية هي ربط المحولات والنسخ العكسي لإنشاء إدراجات cDNA مؤطرة بتسلسل مطلوب لتسلسل الجيل التالي (NGS) باستخدام منصة Illumina. المرحلة الأخيرة من إعداد المكتبة هي رد فعل PCR حيث يتم تضخيم البنى ووضع علامات عليها مع عينة من الباركود المحدد للسماح بتعدد وتسلسل عينات مختلفة على قناة واحدة. قبل التسلسل، يتم تقييم جودة وكمية المكتبات من خلال الحمض النووي عالي الحساسية على رقاقة الكهربائي. يمكن بعد ذلك تجميع مكتبات cDNA ذات المعلمات المناسبة وتسلسلها. يمكن إجراء التسلسل على منصات Illumina مختلفة، مثل MiSeq، NextSeq أو HighSeq، اعتمادا على عدد المكتبات، وطول القراءة المطلوبة وعمق التسلسل. بعد التسلسل، يتم إجراء التحليل المعلوماتي الحيوي.

الشكل 2. إعداد المكتبة. يتضمن إعداد المكتبة إصلاح النهايات وربط المحولات والنسخ العكسي والتضخيم باستخدام ترميز. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
التنميط ريبوسوم هو أسلوب عالمي التي يمكن تعديلها بسهولة وتعديلها وفقا للمسألة العلمية. في الأصل كان يستخدم في الخميرة1، ولكن بعد فترة وجيزة تم تطبيقه على الخلايا البكتيرية21 وكذلك الكائنات الحية نموذج eukaryotic بما في ذلك الماوس10، حمار وحشي22، ذبابة الفاكهة23 و Arabidopsis thaliana24. كما تم استخدامه لدراسة أنواع مختلفة من الريبوسوم: السيتوبلازمية، الميتوكوندريا25،26 وكلوروبلاست27،28. في eukaryotes RIBO-seq عادة ما يتم تكييفها وصقلها للتحقيق في جوانب محددة من الترجمة، بما في ذلك بدء10،11،29،30،31،32، استطالة1،10،11،31،33، ريبوسوم المماطلة33 وتغيير تشكيل33. معظم التعديلات تنطوي على استخدام مثبطات الترجمة المختلفة. في البكتيريا ومع ذلك، كانت الدراسات المماثلة من الصعب إجراء بسبب ندرة مثبطات مع الآلية المطلوبة للعمل34. مثبط الترجمة الأكثر استخداما في البكتيريا هو الكلورامفينيكول (CAM) الذي يرتبط بمركز نقل بيبتيديل (PTC) ويمنع تحديد المواقع الصحيحة لل AMINOACYL-tRNA في الموقع A. ونتيجة لذلك، يمنع CAM تشكيل رابطة الببتيد مما يؤدي إلى اعتقال ريبوسومات ممدود35. أمثلة أخرى من مثبطات الترجمة في البكتيريا هي التتراسيكلين (TET)36، retapamulin (RET)34 و Onc11237 التي استخدمت للتحقيق في مواقع بدء الترجمة. TET، الذي يمنع تسليم الحمض النووي الريبي إلى ريبوسوم عن طريق التداخل المباشر مع حلقة الجذعية anticodon من tRNA في الموقع A، تم تطبيقها أصلا للتحقق من النتائج التي تم الحصول عليها من العلاج CAM لأنها على حد سواء المضادات الحيوية تثبيط ترجمة استطالة38. تم العثور على TET للكشف عن الأولية، ولكن لم يتمكن من الكشف عن TIS36الداخلية. يربط RET في PTC من الريبوسوم البكتيري ، ويمنع تكوين أول رابطة الببتيد عن طريق التدخل في استطالة أميناسيل -tRNA في الموقع A. تطبيق نتائج RET في اعتقال الريبوسومات في كل من الابتدائية وكذلك الداخلية TISS34. Onc112، ببتيد مضاد للميكروبات غني بالبرولين، يربط في نفق الخروج ويمنع ربط أمينواسيل-ترنا في موقع ريبوسومال A. ونتيجة لذلك، يمنع Onc112 مجمعات البدء من دخول مرحلة الاستطالة37.
المعلومات الرئيسية التنميط ريبوسوم يوفر هو كثافة الريبوسومات وموقفها على مرنا. تم تطبيقه بنجاح للتحقيق في التعبير الجيني التفاضلي على مستوى الترجمة في ظروف النمو المختلفة1،6، قياس الكفاءة الترجمية1،38،39 والكشف عن أحداث تنظيم الترجمة مثل التوقف الريبوسومي10. RIBO-seq يسمح أيضا للكشف عن ترجمة ncRNA المشروح، pseudogenes وإطارات القراءة المفتوحة الصغيرة غير معطاة (ORF) مما يؤدي إلى تحديد الجينات الرواية و / أو قصيرة جدا البروتين الترميز10،12،22،30،37. في مثل هذه الحالات، يمكن RIBO-seq صقل وتحسين التعليقات التوضيحية الجينوم. مع حساسيته العالية لتحديد ORFs المترجمة وطبيعتها الكمية ، يمكن أن يكون التنميط الريبوسوم أيضا بمثابة وكيل لتحديد البروتيوم أو في مساعدة دراسات البروتيوميات31،34،39. عن طريق رسم خرائط TIS، التنميط ريبوسوم يكشف N-ميؤوس منها isoforms الموسعة والمقتطعة من البروتينات المعروفة10،32. كما تم تكييف RIBO-seq لدراسة الطي المترجم المشترك للبروتينات14و21و24. تمكن هذه الطريقة من قياس معدلات الاستطالة1،10،39 أو فك سرعة codons الفردية6 وتساعد في تطوير نماذج كمية للترجمة17. طريقة التنميط ريبوسوم هي أيضا قادرة على توفير رؤى ميكانيكية في التوقف ريبوسوم في البكتيريا7،15،17، frameshifting40، وقف كودون قراءة21، إنهاء / إعادة تدوير العيوب41،42 وتشكيل ريبوسومات التغييرات33 في eukaryotes. كما تم تكييف RIBO-seq لدراسة تأثير عوامل محددة عبر المفعول على الترجمة مثلميرناس 6 والبروتينات الرابطة للجيش الملكي النيبالي في eukaryotes16،43. ومع ذلك، من المهم أن نعترف بأن التصميم التجريبي والقرار الذي تم الحصول عليه من ريبو-seq تحديد كمية المعلومات التي يمكن استخلاصها من البيانات الناتجة التسلسل12.
