La technique de profilage des ribosomes (RIBO-seq) a été développée dans le laboratoire de Jonathan Weissman à l’Université de Californie à San Francisco1. En comparaison avec d’autres méthodes utilisées pour étudier l’expression des gènes au niveau translationnel, RIBO-seq se concentre sur chaque ribosome se liant à l’ARNm et fournit des informations sur son emplacement et le nombre relatif de ribosomes sur une transcription. Il permet de surveiller le processus de synthèse des protéines in vivo et peut fournir une résolution et une précision de codon unique permettant la mesure de la densité du ribosome sur les deux, l’ARNm individuel et le long de l’ensemble du transcriptome dans la cellule. A la base de la technique RIBO-seq se trouve le fait que lors de la traduction le ribosome lie la molécule d’ARNm et protège ainsi le fragment enfoui de la transcription d’une digestion ribonucléase. Lors de l’ajout de la ribonucléase, l’ARNm non protégé est digéré et les fragments enfermés par des ribosomes – généralement de ~ 28-30 nt de long – restent intacts. Ces fragments, appelés empreintes ribosomiques (RF), peuvent ensuite être isolés, séquencés et cartographiés sur la transcription dont ils sont issus, ce qui a permis de détecter la position exacte des ribosomes. En fait, la capacité du ribosome à protéger les fragments d’ARNm est utilisée depuis les années 1960 pour étudier les sites ribosomiques de liaison et d’initiation de traduction (TIS)2,3,4. Cependant, avec les progrès de la technologie de séquençage en profondeur, RIBO-seq est devenu une référence pour la surveillance de la traduction5 qui, grâce à l’engagement du ribosome, peut fournir une information à l’échelle du génome sur la synthèse des protéines6. Le profilage du ribosome a comblé le fossé technologique qui existait entre la quantification du transcriptome et le protéome6.
Pour effectuer le profilage du ribosome, nous devons obtenir le lysat cellulaire de l’organisme qui s’était développé dans les conditions étudiées. La perturbation de ces conditions pendant la collecte et la lyse cellulaires peut fournir des données peu fiables. Pour éviter cela, les inhibiteurs de traduction, la récolte rapide et la congélation instantanée dans l’azote liquide sont couramment utilisés. Les cellules peuvent être lysées par broyage cryogénique dans un homogénéisateur mécanique tel qu’un mélangeur7,8 ou un batteur de perles9,et par trituration à travers une pipette10 ou avec une aiguille11. Le tampon de lyse peut être ajouté juste avant ou peu de temps après la pulvérisation des cellules. Dans notre protocole, nous utilisons de l’azote liquide pour pré-refroidir le mortier et le pilon, ainsi que de l’oxyde d’aluminium comme une approche plus douce de la perturbation de la paroi cellulaire bactérienne, ce qui empêche le cisaillement de l’ARN souvent rencontré lorsque des méthodes telles que la sonification sont appliquées. Après pulvérisation, nous ajoutons un tampon de lyse glacé dans le contenu refroidi du mortier. La sélection d’un tampon de lyse approprié est importante pour obtenir la meilleure résolution des empreintes ribosomiques. Étant donné que la force ionique affecte à la fois la taille rf et la précision du cadre de lecture, il est actuellement recommandé d’utiliser des tampons de lyse à faible force ionique et capacité tampon, même s’il apparaît que la composition tampon n’affecte pas l’occupation ribosomique sur les ARNm11,12. Les composants importants du tampon de lyse sont les ions magnésium, dont la présence empêche la dissociation des sous-unités ribosomiques et inhibe les changements conformationnels spontanés dans les ribosomes bactériens11,13. Les ions calcium jouent également un rôle important et sont essentiels pour l’activité de la nucléase micrococcique (MNase) utilisée dans la méthode de profilage des ribosomes bactériens14. L’addition de guanosine 5′-[β,γ-imido]triphosphate (GMP-PNP), un analogue non hydrolysable du GTP, avec le chloramphénicol inhibe la traduction pendant la lyse15.
Lorsque le lysat est obtenu, il est clarifié par centrifugation et divisé en deux parties, chacune pour un RIBO-seq et un séquençage total à haut débit de l’ARNm (RNA-seq) puisqu’ils sont effectués simultanément(figure 1). RNA-seq fournit un point de référence qui permet la comparaison des données de RIBO-seq et RNA-seq lors de l’analyse des données. Le translatome étudié est défini par la normalisation des empreintes ribosomiques à l’abondance de l’ARNm16. Les données de RNA-seq peuvent également aider à identifier le clonage ou le séquençage d’artefacts17.
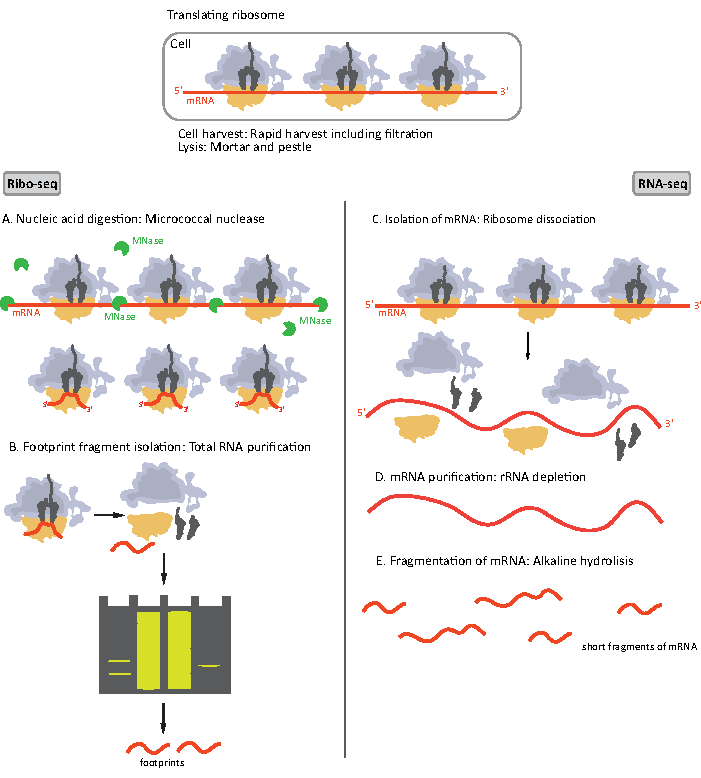
Figure 1. Schémas de préparation d’échantillons d’ARNm pour RIBO-seq et RNA-seq. Pour la préparation de la bibliothèque RIBO-seq, l’ARN est digéré avec de la MNase (A), suivie de la sélection de la taille de RF d’une longueur d’environ 28-30 nt (B); pour l’ARN-seq, l’ARN est isolé (C), appauvri en ARNr (D), et l’ARNm résultant est fragmenté aléatoirement en fragments de longueurs variables (E). Veuillez cliquer ici pour voir une version plus grande de cette figure.
Les étapes initiales de la procédure de préparation de l’échantillon pour RIBO-seq et RNA-seq diffèrent légèrement(Figure 1). Pour le profilage ribosomique, le lysat doit être digéré par une endonucléase spécifique pour dégrader les molécules d’ARNm non protégées par les ribosomes. Dans les protocoles standards, les monosomes obtenus sont récupérés par un coussin de saccharose ultracentrifugation ou un gradient de saccharose d’ultracentrifugation8,14. Dans cet article, nous montrons que cette étape n’est pas nécessaire pour isoler le RF requis pour le RIBO-seq dans les bactéries, de même pour les cellules eucaryotes18,et que la sélection de taille des fragments d’ARNm de longueur appropriée à partir du gel de polyacrylamide est suffisante.
Pour l’ARN-seq, l’ARNm est obtenu par épuisement de l’ARNr à partir de l’ARN total – les molécules d’ARNr s’hybrident aux sondes oligonucléotidiques biotinylées qui se lient aux billes magnétiques recouvertes de streptavidine. Les complexes ARNr-oligonucléotide-billes sont ensuite retirés de l’échantillon avec un aimant aboutissant à un arnr appauvri échantillon19,20. Les molécules d’ARNm purifiées sont ensuite fragmentées aléatoirement par hydrolyse alcaline. Les fragments d’ARNm obtenus ainsi que les empreintes ribosomiques sont convertis en bibliothèques d’ADNc et préparés pour le séquençage en profondeur (Figure 2). Cela implique la réparation des extrémités nécessaires après hydrolyse alcaline (pour l’ARNm) et la digestion enzymatique (pour RF): déphosphorylation des extrémités 3′ suivie d’une phosphorylation des extrémités 5′. Les étapes suivantes sont la ligature des adaptateurs et la transcription inverse pour créer des inserts d’ADNc encadrés par des séquences requises pour le séquençage de nouvelle génération (NGS) à l’aide de la plate-forme Illumina. La dernière phase de la préparation de la bibliothèque est une réaction pcr dans laquelle les constructions sont amplifiées et étiquetées avec des codes à barres spécifiques à l’échantillon pour permettre le multiplexage et le séquençage de divers échantillons sur un canal. Avant le séquençage, la qualité et la quantité des bibliothèques sont évaluées par électrophorèse sur puce à haute sensibilité de l’ADN. Les bibliothèques d’ADNc avec les paramètres appropriés peuvent ensuite être regroupées et séquencées. Le séquençage peut être effectué sur différentes plates-formes Illumina, telles que MiSeq, NextSeq ou HighSeq, en fonction du nombre de bibliothèques, de la longueur de lecture requise et de la profondeur de séquençage. Après le séquençage, l’analyse bioinformatique est effectuée.

Figure 2. Préparation de la bibliothèque. La préparation de la bibliothèque comprend la réparation des extrémités, la ligature des adaptateurs, la transcription inverse et l’amplification avec codage à barres. Veuillez cliquer ici pour voir une version plus grande de cette figure.
Le profilage des ribosomes est une méthode universelle qui peut être facilement modifiée et ajustée en fonction de la question scientifique. À l’origine, il a été utilisé dans la levure1, mais peu de temps après, il a été appliqué sur les cellules bactériennes21 ainsi que sur des organismes modèles eucaryotes, y compris la souris10,le poisson zèbre22,la mouche des fruits23 et Arabidopsis thaliana24. Il a également été utilisé pour étudier différents types de ribosomes: cytoplasmique, mitochondrial25,26 et chloroplaste27,28. Chez les eucaryotes, ribo-seq est couramment adapté et affiné pour étudier des aspects spécifiques de la traduction, y compris l’initiation10,11,29,30,31,32,l’allongement1,10,11,31,33,le ribosome calant33 et le changement de conformation33. La plupart des modifications impliquent l’utilisation de différents inhibiteurs de traduction. Chez les bactéries cependant, des études analogues ont été difficiles à mener en raison de la rareté des inhibiteurs avec le mécanisme d’action requis34. L’inhibiteur de traduction le plus couramment utilisé chez les bactéries est le chloramphénicol (CAM) qui se lie au centre de la peptidyl transférase (PTC) et empêche le positionnement correct de l’aminoacyl-ARNt dans le site A. En conséquence, cam empêche la formation d’une liaison peptidique qui conduit à arrêter les ribosomes allongés35. D’autres exemples d’inhibiteurs de traduction chez les bactéries sont la tétracycline (TET)36,la retapamuline (RET)34 et l’Onc11237 qui ont été utilisés pour étudier les sites d’initiation de la traduction. Le TET, qui empêche la livraison de tRNA au ribosome en chevauchant directement avec l’anticodon tige-boucle du tRNA au A-site, a été à l’origine appliqué pour vérifier les résultats obtenus du traitement de CAM puisqu’ils sont les deux antibiotiques inhibant l’élongation de traduction38. TET s’est avéré pour détecter tis primaire, cependant n’a pas pu indiquer TISinterne 36. RET se lie au PTC du ribosome bactérien, et empêche la formation de la première liaison peptidique en interférant avec un aminoacyl-ARNt allongé dans le site A. L’application de RET entraîne l’arrêt des ribosomes à la fois primaires et internesTIS 34. Onc112, un peptide antimicrobien riche en proline, se lie dans le tunnel de sortie et bloque la liaison aminoacyl-ARNt dans le site A ribosomique. En conséquence, Onc112 empêche les complexes d’initiation d’entrer dans la phase d’allongement37.
La principale information que fournit le profilage des ribosomes est la densité des ribosomes et leur position sur l’ARNm. Il a été appliqué avec succès pour étudier l’expression différentielle des gènes au niveau de la traduction dans diverses conditions de croissance1,6,mesurer l’efficacité translationnelle1,38,39 et détecter les événements de régulation de la traduction tels que la suspension ribosomique10. Ribo-seq permet également de découvrir la traduction d’ARNc annotés, de pseudogènes et de petits cadres de lecture ouverts non annotés (ORF) conduisant à l’identification de gènes codant des protéines nouvelles et/ou très courtes10,12,22,30,37. Dans de tels cas, RIBO-seq peut affiner et améliorer l’annotation du génome. Avec sa grande sensibilité pour l’identification des ORF traduits et sa nature quantitative, le profilage des ribosomes peut également servir d’indicateur pour la détermination du protéome ou pour faciliter les études protéomiques31,34,39. En cartographiant le TIS, le profilage des ribosomes révèle des isoformes N-terminalement étendues et tronquées de protéines connues10,32. Ribo-seq a également été adapté pour étudier le repliement co-traductionnel des protéines14,21,24. Cette méthode permet de mesurer des vitesses d’allongement1,10,39 ou de décoder des codons individuels6 et aide à développer des modèles quantitatifs de translation17. La méthode de profilage du ribosome est également capable de fournir des informations mécanistes sur le ribosome en pause chez les bactéries7,15,17,le déphasage40,la lecture stop-codon21,les défauts de terminaison/recyclage41,42 et les changements de conformation ribosomique33 chez les eucaryotes. Ribo-seq a également été adapté pour examiner l’impact de facteurs trans-agissants spécifiques sur la traduction tels que les miARN6 et les protéines de liaison à l’ARN chez les eucaryotes16,43. Cependant, il est important de reconnaître que le plan expérimental et la résolution obtenue de RIBO-seq déterminent la quantité d’informations qui peuvent être extraites des données de séquençage résultantes12.
