リボソームプロファイリング技術(RIBO-seq)は、カリフォルニア大学サンフランシスコ校のジョナサン・ワイスマンの研究室で開発されました。RIBO-seqは、トランスレーショナルレベルで遺伝子発現を研究するために使用される他の方法と比較して、mRNAに結合する各リボソームに焦点を当て、その位置およびトランスクリプト上のリボソームの相対的な数に関する情報を提供します。それは、生体内でのタンパク質合成のプロセスを監視することを可能にし、細胞内の個々のmRNAおよび全転写体に沿って両方のリボソーム密度の測定を可能にする単一のコドンの分解能および正確さを提供することができる。RIBO-seq技術の基礎には、翻訳中にリボソームがmRNA分子に結合し、リボヌクレアーゼ消化から転写物の埋もれた断片を保護するという事実があります。リボヌクレアーゼを添加すると、保護されていないmRNAが消化され、リボソーム(通常〜28〜30 ntの長さ)で囲まれた断片はそのまま残ります。リボソームフットプリント(RF)と呼ばれるこれらの断片は、リボソームの正確な位置を検出することから生まれたトランスクリプトに分離、配列決定、マッピングすることができます。実際、mRNA断片を保護するリボソーム能力は、1960年代からリボソーム結合および翻訳開始部位(TIS)2、3、4を研究するために使用されてきた。しかし、深いシーケンシング技術の進歩に伴い、RIBO-seqは、リボソーム関与を通じて、タンパク質合成に関するゲノム全体の情報を提供することができる翻訳モニタリング5のゴールドスタンダードとなっている。リボソームプロファイリングは、トランスクリプトームとプロテオーム6を定量化する間に存在する技術的なギャップを埋めた。
リボソームプロファイリングを行うには、調査対象条件下で増殖した生物の細胞リセートを取得する必要があります。細胞採取およびリシス中にこれらの条件を中断すると、信頼性の低いデータが得られ得る。これを防ぐために、翻訳阻害剤、液体窒素中の急速な収穫およびフラッシュ凍結が一般的に使用されている。細胞は、ミキサーミル7、8またはビーズビーター9のような機械的ホモジナイザーでの極低温粉砕によって、ピペット10または針11でトリチュレーションすることによって、凍結粉砕することができる。細胞の粉砕の直前または直後に、分解バッファーを追加できます。私たちのプロトコルでは、乳鉢や害虫を予冷するために液体窒素を使用し、細菌細胞壁の破壊に対する優しいアプローチとして酸化アルミニウムを使用し、ソニフィケーションなどの方法が適用されたときにしばしば遭遇するRNAせん断を防ぎます。粉砕後、乳鉢の冷却された内容物に氷冷のライシスバッファーを追加します。リボソームフットプリントの最適な解像度を得るためには、適切なリシスバッファーの選択が重要です。イオン強度はRFサイズと読み取りフレーム精度の両方に影響を与えるので、現在では、バッファー組成がmRNA1,12のリボソーム占有率に影響を与えないと思われるとしても、イオン強度と緩衝能の低いリシスバッファを使用することが推奨されている。このリシスバッファーの重要な成分はマグネシウムイオンであり、その存在はリボソームサブユニットの解離を防止し、細菌リボソーム11,13における自然な立体構造変化を阻害する。カルシウムイオンもまた重要な役割を果たし、細菌性リボソームプロファイリング法14で用いられる微小球核化(MNase)の活性に必須である。グアノシン5′の添加-[β,γ-imido]三リン酸(GMP-PNP)は、GTPの非加水分解可能なアナログであり、クロラムフェニコールと共に、リシス15の間の翻訳を阻害する。
ライセートが得られると、遠心分離によって明確化され、RIBO-seqと高スループットの総mRNAシーケンシング(RNA-seq)が同時に行われるため、それぞれ2つの部分に分けられる(図1)。RNA-seqは、データ分析中にRIBO-seqとRNA-seqの両方からのデータを比較できる基準点を提供します。調査した翻訳は、リボソームフットプリントをmRNAの存在量16に正規化することによって定義される。RNA-seq からのデータは、クローニングまたはシーケンシングアーティファクト17を識別するのにも役立ちます。
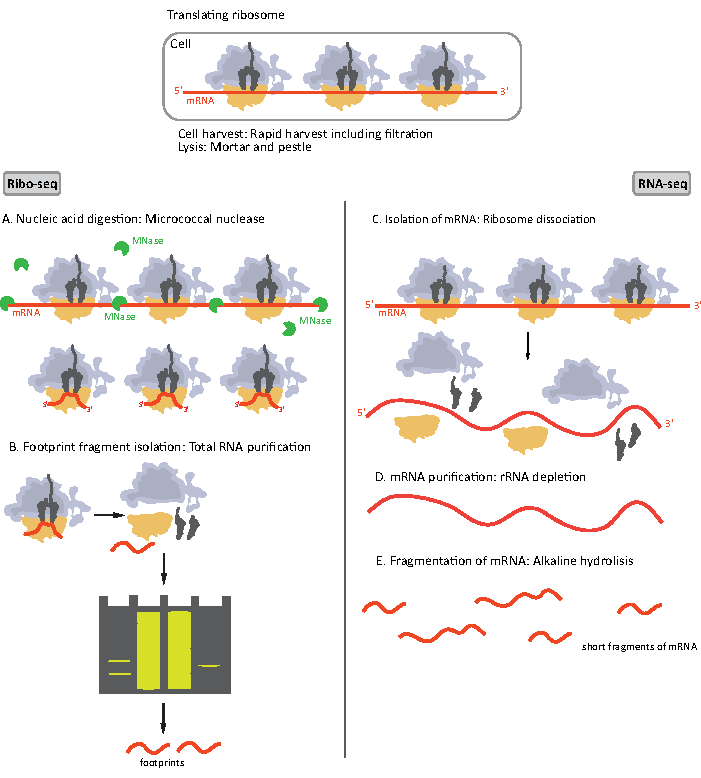
図 1.RIBO-セクおよびRNA-セクに対するmRNAサンプル調製物の概略図。 RIBO-seqライブラリ調製の場合、RNAはMNase(A)で消化され、続いて〜28〜30 ntの長さ(B)のRFのサイズ選択が続きます。RNA-セクRNAが単離(C)であるため、rRNA(D)の枯渇、および得られたmRNAは、様々な長さの断片(E)にランダムに断片化される。 この図の大きなバージョンを表示するには、ここをクリックしてください。
RIBO-seqとRNA-seqのサンプル調製手順の初期ステップは若干異なる(図1)。リボソームプロファイリングの場合、リソソームによって保護されていないmRNA分子を分解するために、特定のエンドヌクレアーゼによってリセートを消化する必要があります。標準的なプロトコルにおいて、得られたモノソームは、ショ糖クッション超遠心分離またはショ糖勾配超遠心8、14によって回収される。本稿では、このステップは、真核生物細胞18に対しても同様に、細菌中のRIBO-seqに必要なRFを分離する必要はないことを示し、ポリアクリルアミドゲルからの適切な長さのmRNA断片のサイズ選択が十分であることを示す。
RNA-seqの場合、mRNAは、全RNA-rRNA分子からストレプトアビジンコーティングされた磁気ビーズに結合するビオチン化オリゴヌクレオチドプローブにハイブリダイズする全RNAからrRNAの枯渇によって得られる。次いで、rRNAオリゴヌクレオチドビーズ複合体を磁石でサンプルから除去し、rRNAが枯渇したサンプル19,20を得る。精製されたmRNA分子は、次いでアルカリ加水分解によってランダムに断片化される。得られたmRNAの断片とリボソーム足跡はcDNAライブラリに変換され、深いシーケンシングのために準備される(図2)。これは、アルカリ加水分解(mRNAの場合)と酵素消化(RF用)の後に必要な修復を終了することを含む:3’末端の脱リン酸化後に5’末端のリン酸化が続く。次のステップは、アダプタライゲーションと逆転写を行い、Illuminaプラットフォームを使用して次世代シーケンシング(NGS)に必要な配列でフレーム化されたcDNAインサートを作成します。ライブラリー調製の最後の段階は、構成体を増幅し、サンプル固有のバーコードでラベル付けし、1つのチャネルでさまざまなサンプルを多重化およびシーケンス処理できるようにする PCR 反応です。シーケンシングの前に、ライブラリの質と量は高感度DNAオンチップ電気泳動によって評価されます。適切なパラメータを持つcDNAライブラリをプールし、シーケンスを作成できます。シーケンスは、ライブラリの数、必要な読み取り長さ、およびシーケンス深度に応じて、MiSeq、NextSeq、HighSeqなどのさまざまなイルミナプラットフォームで実行できます。シーケンシング後、バイオインフォマティクス解析が行われる。

図 2.図書館の準備。 ライブラリの調製は、エンド修復、アダプターライゲーション、逆転写およびバーコードによる増幅を含む。 この図の大きなバージョンを表示するには、ここをクリックしてください。
リボソームプロファイリングは、科学的な質問に従って簡単に修正および調整することができる普遍的な方法です。もともとは酵母1で使用されていたが、マウス10、ゼブラフィッシュ22、フルーツフライ23及びシロイヌナズナ24を含む真核生物と同様に細菌細胞21に適用された直後に。また、細胞質、ミトコンドリア25、26、葉葉芽細胞27、28の異なるリボソームタイプの研究にも使用されました。真核生物では、RIBO-seqは、一般的に、開始10、11、29、30、31、32、伸び1、10、11、31、33、リボソーム失速33およびコンフォメーション変化33を含む翻訳の特定の側面を調査するために適応および精製される。ほとんどの修飾は、異なる翻訳阻害剤の使用を含む。しかし、細菌では、作用機序34を有する阻害剤の貧弱性のために類似の研究が行いにくい。細菌の中で最も一般的に使用される翻訳阻害剤は、ペプチジルトランスファーゼセンター(PTC)に結合し、Aサイトにおけるアミノアシル-tRNAの正しい位置を防ぐクロラムフェニコール(CAM)です。その結果、CAMは、伸びるリボソーム35を阻止することにつながるペプチド結合の形成を防止する。細菌における翻訳阻害剤の他の例としては、テトラサイクリン(TET)36、リタパミュリン(RET)34およびOnc11237があり、翻訳開始部位の調査に使用されている。TETは、A-サイトでのtRNAの抗コドンステムループと直接重なることによってリボソームへのtRNA送達を防止し、これらは共に抗生物質が翻訳伸長38を阻害するのでCAM治療から得られた結果を検証するために適用された。TET はプライマリ TIS を検出することが判明したが、内部 TIS36を明らかにすることができなかった。RETは細菌リボソームのPTCに結合し、A部位におけるアミノアシル-tRNAを細長く干渉させることにより第1のペプチド結合の形成を防止する。RETを適用すると、プライマリと内部のTIS34の両方でリボソームが逮捕されます。Onc112は、プロリンが豊富な抗菌ペプチドであり、出口トンネル内で結合し、リボソームA部位におけるアミノアシル-tRNA結合をブロックする。その結果、Onc112は開始複合体が伸びフェーズ37に入るのを防ぎます。
リボソームプロファイリングが提供する主な情報は、リボソーム密度およびmRNA上でのそれらの位置である。様々な成長条件1,6の翻訳レベルでの遺伝子発現の差動を調べ、翻訳効率1、38、39を測定し、リボソーム休止などの翻訳調節事象を検出するために応用に成功した。RIBO-seqはまた、新しいおよび/または非常に短いタンパク質コード遺伝子10、12、22、30、37の同定につながる、アノト化ncRNA、擬似遺伝子および無公認の小さなオープンリーディングフレーム(ORF)の翻訳を明らかにすることを可能にする。このような場合、RIBO-seqはゲノムアノテーションを微調整し、改善することができます。翻訳されたORFの同定とその定量的性質に対する高感度を有するリボソームプロファイリングは、プロテオームの決定の代理としても、またはプロテオミクス研究31、34、39を補助する役割を果たすことができる。TISをマッピングすることにより、リボソームプロファイリングは、既知のタンパク質10,32のN末端拡張および切り捨てられたアイソフォームを明らかにする。RIBO-seqはまた、タンパク質14、21、24の共翻訳折りたたみを研究するために適応された。この方法は、個々のコドン6の伸び速度1、10、39またはデコード速度の測定を可能にし、翻訳17の定量モデルの開発に役立ちます。リボソームプロファイリング法は、細菌7、15、17、フレームシフト40、ストップコドン読み出し21、終了/リサイクル欠陥41、42およびリボソーム立体構造変化33の細菌におけるリボソーム休止に対する機械学的洞察を提供することもできる。RIBO-seqはまた、真核生物におけるmiRNA6およびRNA結合タンパク質などの翻訳に対する特異的トランス作用因子の影響を調べるためにも適応された。しかしながら、RIBO-seqの実験計画および得られた解像度が、得られたシーケンシングデータ12から抽出できる情報量を決定することを認めることが重要である。
