La tecnica di profilazione ribosoma (RIBO-seq) è stata sviluppata nel laboratorio di Jonathan Weissman presso l’Università della California, San Francisco1. Rispetto ad altri metodi usati per studiare l’espressione genica a livello traslazionale, RIBO-seq si concentra su ogni legame ribosoma all’mRNA e fornisce informazioni sulla sua posizione e sul numero relativo di ribosomi su una trascrizione. Consente di monitorare il processo di sintesi proteica in vivo e può fornire una risoluzione e una precisione del codone singolo consentendo la misurazione della densità ribosoma su entrambi, il singolo mRNA e lungo l’intero trascrittame nella cellula. Alla base della tecnica RIBO-seq si trova il fatto che durante la traduzione il ribosoma lega la molecola di mRNA e quindi protegge il frammento sepolto della trascrizione da una digestione della ribonucleasi. Dopo l’aggiunta della ribonucleasi, l’mRNA non protetto viene digerito e i frammenti racchiusi da ribosomi – tipicamente lunghi ~28-30 nt – rimangono intatti. Questi frammenti, chiamati impronte ribosomiali (RF), possono quindi essere isolati, sequenziati e mappati sulla trascrizione da cui hanno avuto origine con conseguente rilevamento della posizione esatta dei ribosomi. Infatti, la capacità ribosoma di proteggere i frammenti di mRNA è stata utilizzata dagli anni ’60 per studiare i siti di rilegatura ribosomiale e iniziazione alla traduzione (TIS)2,3,4. Tuttavia, con l’avanzamento della tecnologia di sequenziamento profondo, RIBO-seq è diventato un gold standard per il monitoraggio dellatraduzione 5 che, attraverso l’impegno ribosoma, può fornire informazioni a livello genomico sulla sintesi proteica6. La profilazione ribosoma ha colmato il divario tecnologico esistente tra la quantificazione del trascrittame e il proteoma6.
Per condurre la profilazione ribosoma è necessario ottenere il lisato cellulare dell’organismo che era cresciuto nelle condizioni studiate. Interrompere queste condizioni durante la raccolta e lalisi cellulare può fornire dati inaffidabili. Per evitare ciò, vengono comunemente utilizzati inibitori della traduzione, raccolta rapida e congelamento lampo nell’azoto liquido. Le cellule possono essere lisciviate mediante macinazione criogenica in un omogeneizzatore meccanico come un miscelatore7,8 o un battitore diperline 9e per triturazione attraverso una pipetta10 o con un ago11. Il tampone di lysis può essere aggiunto poco prima o poco dopo la polverizzazione delle cellule. Nel nostro protocollo utilizziamo azoto liquido per preraffreddare malta e pestello, così come l’ossido di alluminio come approccio più delicato all’interruzione della parete cellulare batterica, che impedisce la tosatura dell’RNA spesso riscontrata quando vengono applicati metodi come la sonificazione. Dopo la polverizzazione, aggiungiamo un tampone di lisi ghiacciata nel contenuto raffreddato della malta. La selezione di un buffer di lisi appropriato è importante per ottenere la migliore risoluzione delle impronte ribosomiali. Poiché la forza ionica influisce sia sulla dimensione RF che sulla precisione del fotogramma di lettura, si consiglia attualmente di utilizzare buffer di lisi con bassa forza ionica e capacità tampone, anche se sembra che la composizione tampone non influenzi l’occupazione ribosomiale su mRNA11,12. Componenti importanti del tampone di lisi sono gli ioni di magnesio, la cui presenza previene la dissociazione delle subunità ribosomiali e inibisce i cambiamenti conformazionali spontanei nei ribosomibatterici 11,13. Anche gli ioni di calcio svolgono un ruolo significativo e sono essenziali per l’attività della nucleasi micrococcica (MNasi) utilizzata nel metodo di profilazione ribosomabatterica 14. L’aggiunta di guanosina 5′-[β,γ-imido]trifosfato (GMP-PNP), un analogo non idrolabile di GTP, insieme al cloramfenicolo inibisce la traduzione durante la lisi15.
Quando il lisato viene ottenuto, viene chiarito per centrifugazione e diviso in due porzioni, ognuna per un RIBO-seq e un sequenziamento totale dell’mRNA ad alta produttività (RNA-seq) poiché vengono eseguite simultaneamente(Figura 1). RNA-seq fornisce un punto di riferimento che consente il confronto dei dati sia ribo-seq che RNA-seq durante l’analisi dei dati. Il translatoma studiato è definito dalla normalizzazione delle impronte ribosomiche all’abbondanza di mRNA16. I dati di RNA-seq possono anche aiutare a identificare la clonazione o sequenziare gliartefatti 17.
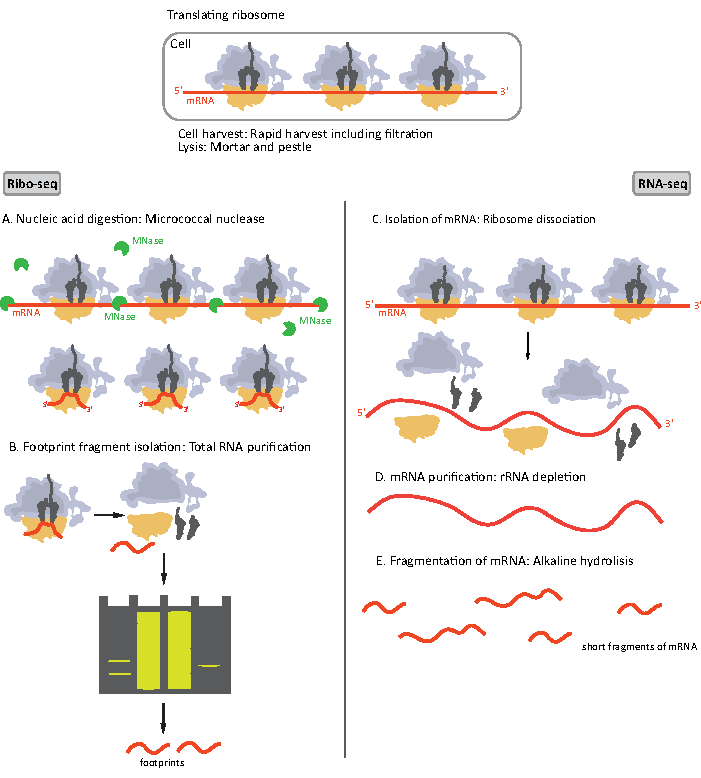
Figura 1. Schemi di preparazione del campione di mRNA per RIBO-seq e RNA-seq. Per la preparazione della libreria RIBO-seq, l’RNA viene digerito con MNasi (A), seguito dalla selezione delle dimensioni di RF di ~28-30 nt di lunghezza (B); per l’RNA RNA-seq è isolato (C), impoverito di rRNA (D), e l’mRNA risultante è frammentato casualmente in frammenti di varie lunghezze (E). Clicca qui per visualizzare una versione più grande di questa figura.
I primi passi della procedura di preparazione del campione per RIBO-seq e RNA-seq differiscono leggermente (Figura 1). Per la profilazione ribosomiale, il lisato deve essere digerito da una specifica endonucleasi per degradare le molecole di mRNA non protette dai ribosomi. Nei protocolli standard, i monosomi ottenuti vengono recuperati da un cuscino di saccarosio ultracentrifugato o da un gradiente di saccarosio ultracentrifugazione8,14. In questo articolo, mostriamo che questo passaggio non è necessario per isolare RF richiesto per il RIBO-seq nei batteri, allo stesso modo per le cellule eucariotiche18e che la selezione delle dimensioni dei frammenti di mRNA di lunghezza appropriata dal gel di poliacrilammide è sufficiente.
Per l’RNA-seq, l’mRNA è ottenuto dall’esaurimento dell’rRNA dalle molecole totali di RNA – rRNA che si ibridano alle sonde oligonucleotidi biotinylate che si legano alle perline magnetiche rivestite di streptavidina. I complessi rRNA-oligonucleotide-perline vengono quindi rimossi dal campione con un magnete con conseguente campionamento impoverito di rRNA19,20. Le molecole di mRNA purificate vengono quindi frammentate casualmente per idrolisi alcalina. I frammenti ottenuti di mRNA e le impronte ribosomiali vengono convertiti in librerie cDNA e preparati per il sequenziamento profondo (Figura 2). Ciò comporta la riparazione delle estremità necessaria dopo l’idrolisi alcalina (per mRNA) e la digestione enzimatica (per RF): defosforilazione di 3′ estremità seguita da fosforilazione di 5′ estremità. I passaggi successivi sono la legatura degli adattatori e la trascrizione inversa per creare inserti cDNA incorniciati da sequenze necessarie per il sequenziamento di nuova generazione (NGS) utilizzando la piattaforma Illumina. L’ultima fase della preparazione della libreria è una reazione PCR in cui i costrutti vengono amplificati ed etichettati con codici a barre specifici del campione per consentire il multiplexing e il sequenziamento di vari campioni su un canale. Prima del sequenziamento, la qualità e la quantità delle librerie sono valutate dall’elettroforesi su chip del DNA ad alta sensibilità. Le librerie cDNA con parametri appropriati possono quindi essere raggruppate e sequenziate. Il sequenziamento può essere eseguito su diverse piattaforme Illumina, come MiSeq, NextSeq o HighSeq, a seconda del numero di librerie, della lunghezza di lettura e della profondità di sequenziamento richieste. Dopo il sequenziamento, viene eseguita l’analisi bioinformatica.

Figura 2. Preparazione della biblioteca. La preparazione della libreria include la riparazione delle estremità, la legatura degli adattatori, la trascrizione inversa e l’amplificazione con codifica a barre. Clicca qui per visualizzare una versione più grande di questa figura.
La profilazione ribosoma è un metodo universale che può essere facilmente modificato e regolato in base alla questione scientifica. Originariamente è stato utilizzato nellievito 1,ma poco dopo è stato applicato alle cellulebatteriche 21 così come agli organismi modello eucariotico tra cui topo10,zebrafish22,mosca dellafrutta 23 e Arabidopsis thaliana24. È stato anche usato per studiare diversi tipi di ribosomi: citoplasmatico, mitocondriale25,26 e cloroplasto27,28. Negli eucarioti RIBO-seq è comunemente adattato e raffinato per indagare aspetti specifici della traduzione, tra cuiiniziazione 10,11,29,30,31,32,allungamento 1,10,11,31,33,ribosoma stallo33 e modifica conformazione33. La maggior parte delle modifiche comporta l’uso di diversi inibitori della traduzione. Nei batteri, tuttavia, studi analoghi sono stati difficili da condurre a causa della scarsità di inibitori con il meccanismo d’azionerichiesto 34. L’inibitore della traduzione più comunemente usato nei batteri è il cloramfenicolo (CAM) che si lega al centro peptidil transferasi (PTC) e impedisce il corretto posizionamento dell’amminoacil-tRNA nel sito A. Di conseguenza, CAM impedisce la formazione di un legame peptidico che porta ad arrestare i ribosomi allunganti35. Altri esempi di inibitori della traduzione nei batteri sono la tetraciclina (TET)36, la retapamulina (RET)34 e l’Onc11237 che sono stati utilizzati per studiare i siti di iniziazione alla traduzione. TET, che impedisce la consegna di tRNA al ribosoma sovrapponendosi direttamente con l’anello staminali anticodonte del tRNA nel sito A, è stato originariamente applicato per verificare i risultati ottenuti dal trattamento CAM poiché sono entrambi antibiotici che inibiscono l’allungamento dellatraslazione 38. Tet è stato trovato per rilevare tis primario, tuttavia non è stato in grado di rivelare tis36 interno. Il RET si lega nel PTC del ribosoma batterico e previene la formazione del primo legame peptidico interferendo con un aminoacil-tRNA allungatore nel sito A. L’applicazione del RET comporta l’arresto dei ribosomi sia al TISs34primario che a quello interno. Onc112, un peptide antimicrobico ricco di prolina, si lega nel tunnel di uscita e blocca il legame amminoacil-tRNA nel sito ribosomiale A. Di conseguenza, Onc112 impedisce ai complessi di iniziazione di entrare nella fase diallungamento 37.
La principale profilazione ribosoma delle informazioni fornisce la densità dei ribosomi e la loro posizione sull’mRNA. È stato applicato con successo per studiare l’espressione genica differenziale a livello di traduzione in varie condizioni dicrescita 1,6,misurare l’efficienza traslazionale1,38,39 e rilevare eventi di regolazione della traduzione come la pausa ribosomiale10. RIBO-seq consente inoltre di scoprire la traduzione di ncRNA annotato, pseudogeni e piccoli telai di lettura aperti senza preavviso (ORF) che portano all’identificazione di geni di codifica proteica nuovi e/o molto brevi10,12,22,30,37. In questi casi, RIBO-seq può perfezionare e migliorare l’annotazione del genoma. Con la sua elevata sensibilità per l’identificazione degli ORF tradotti e la sua natura quantitativa, la profilazione ribosoma può anche servire da proxy per la determinazione del proteoma o per aiutare gli studi di proteomica31,34,39. Mappando TIS, la profilazione ribosoma rivela isoforme estese e troncate N-terminale di proteinenote 10,32. RIBO-seq è stato anche adattato per studiare il ripiegamento co-traslazionaledelle proteine 14,21,24. Questo metodo consente di misurare i tassi diallungamento 1,10,39 o velocità di decodifica dei singolicodoni 6 e aiuta a sviluppare modelli quantitativi ditraduzione 17. Il metodo di profilazione ribosoma è anche in grado di fornire approfondimenti meccanicistici sulla pausa ribosoma neibatteri 7,15,17,frameshifting40,stop-codon readthrough21,difetti di terminazione /riciclaggio41,42 e modifiche alla conformazione ribosomiale33 negli eucarioti. RIBO-seq è stato anche adattato per esaminare l’impatto di specifici fattori trans-acting sulla traduzione come le miRNA6 e le proteine leganti l’RNA negli eucarioti16,43. Tuttavia, è importante riconoscere che il progetto sperimentale e la risoluzione ottenuta di RIBO-seq determinano la quantità di informazioni che possono essere estratte dai dati di sequenziamentorisultanti 12.
