DNA-methylatie is een belangrijk epigenetisch merkteken dat genexpressie en chromatinestructuur reguleert. Methylering treedt voornamelijk op in CpG-dinucleotiden – cytosine gevolgd door guanosine; de methylgroep wordt toegevoegd aan de 5-positie van cytosine. Correcte DNA-methylatiepatronen, en dus de juiste genexpressie, zijn nodig voor een geschikte cellulaire ontwikkeling en functie. Veel ziektetoestanden zijn in verband gebracht met veranderingen in het normale methylatiepatroon 1,2,3. Er is bijvoorbeeld een verband tussen kankerinitiatie en progressie en veranderingen in het DNA-methylatiepatroon. Typisch, kankercellen vertonen lagere algemene niveaus van methylcytosine, die bijdraagt aan genoominstabiliteit. Tegelijkertijd is de methylcytosine die aanwezig is in het genoom geconcentreerd in de promotorregio’s van tumorsuppressorgenen, wat leidt tot gen-uitschakeling van deze belangrijke eiwitten. Met name epigenetische veranderingen zijn dynamisch en omkeerbaar, in tegenstelling tot de DNA-mutaties geassocieerd met tumorigenese. Dit heeft de eiwitten die betrokken zijn bij epigenetische genregulatie interessante medicijndoelen 2,4 gemaakt.
DNA-methyltransferasen (DNT’s) zijn de eiwitten die verantwoordelijk zijn voor het genereren en onderhouden van DNA-methylatiepatronen. Drie katalytisch actieve isozymen, DNMT1, DNMT3a en DNMT3b, bestaan bij mensen. Tijdens de ontwikkeling en differentiatie stellen de de novo methyltransferasen, DNMT3a en DNMT3b, methylatiepatronen vast. Beide enzymen kunnen het katalytisch inactieve DNMT3L-eiwit binden om complexen te vormen die verhoogde activiteit vertonen 1,5. Na celdeling bevatten dochtercellen hemimethylated DNA – DNA dat methylcytosine bevat in slechts één streng van de duplex – omdat het nieuw gesynthetiseerde DNA verstoken is van methylatiemarkeringen. De belangrijkste functie van DNMT1 is om dit gehemimethyleerde DNA te methyleren, waardoor het volledige methylatiepatroonwordt hersteld 1,5.
Verbanden tussen DNMT-activiteit en kanker zijn goed vastgesteld. Overexpressie van DNMT1, hetzij door transcriptionele of posttranslationele mechanismen, is een gevolg van verschillende veel voorkomende oncogene routes 6,7,8,9. Genetische benaderingen voor lagere DNMT1-activiteit met behulp van hypomorfe allelen resulteren in verminderde tumorvorming bij Apc(Min)-muizen10. Antisense oligonucleotiden die DNMT1 afstoten remmen neoplasie in celkweek en muistumormodellen 11,12. Het remmen van DNMT1-activiteit lijkt dus een veelbelovende benadering van kankertherapie. De rollen die de DNMT3-isozymen spelen, zijn echter niet zo eenvoudig. DNMT3a mutaties worden gevonden bij acute myeloïde leukemie13 en myelodysplastisch syndroom14. Van ten minste één van de geïdentificeerde mutaties is aangetoond dat het de DNA-methylatieactiviteit van het enzym15 vermindert. DNMT3b is echter overexpressie bij borstkanker16 en colorectale kanker17. Omdat de verschillende DNMT-isozymen verschillende rollen spelen in carcinogenese, zal het identificeren van isozymspecifieke remmers van cruciaal belang zijn. Niet alleen zullen deze verbindingen nuttig zijn voor de ontwikkeling van therapeutica, maar isozymspecifieke remmers zouden ook een waardevol hulpmiddel zijn om de rol van elk DNMT-isozym in de kankeretiologie te ontleden.
Verschillende DNMT-remmers zijn gemeld in de literatuur. Bekende DNMT-remmers kunnen worden onderverdeeld in twee klassen: nucleoside en niet-nucleoside. Nucleosideremmers zijn meestal cytidine-analogen. Deze verbindingen zijn opgenomen in DNA en vangen covalent DNMT’s. 5-azacytidine en 5-aza-2′-deoxycytidine zijn goedgekeurd voor de behandeling van myelodysplastisch syndroom en acute myeloïde leukemie 4,18. De hoge toxiciteit, lage biologische beschikbaarheid en chemische instabiliteit van deze verbindingen leveren problemen op. Lopend onderzoek is naar de werkzaamheid van de volgende generatie nucleosideremmers; SGI-110, afgeleid van 5-aza-2′-deoxycytidine, is een voorbeeld19,20. Nucleosideremmers zijn niet isozymspecifiek en zullen elk DNMT-isozym dat wordt aangetroffen inactiveren. Daarom resulteert behandeling met een nucleoside-demethylerend middel in de uitputting van alle DNMT-isozymen 4,18. Niet-nucleosideremmers hoeven niet in het DNA te worden opgenomen om hun remmende effecten uit te oefenen. In plaats daarvan binden deze moleculen zich rechtstreeks aan DNT’s, waardoor de mogelijkheid voor isozymspecifieke remming wordt geïntroduceerd. Tot op heden zijn verschillende niet-nucleosideremmers ontdekt, waaronder SGI-102721, hydralazine22, procaïnamide23, RG108 en derivaten24, en natuurlijke producten, (−)-epigallocatechine 3-gallaat (EGCG)25 en laccaïnezuur A26,27. De meeste niet-nucleosideremmers die tot nu toe zijn ontdekt, zijn niet isozymselectief of vertonen zwakke voorkeuren voor één DNMT-isozym. Bovendien moet de potentie van deze moleculen worden verbeterd, vooral in cellen 4,18. Er is dus behoefte aan het ontdekken of ontwikkelen van krachtigere, isozymselectieve DNMT-remmers.
Een hindernis voor het ontdekken van nieuwe kleine molecuulremmers van DNT’s zijn de moeizame testen die traditioneel worden gebruikt om DNMT-activiteitte onderzoeken 28. Assays zijn meestal discontinu met meerdere stappen. De enzymatische activiteit van DNMT’s wordt nog steeds routinematig getest met behulp van radioactief S-adenosyl methionine (SAM)29,30,31,32,33,34. Er zijn ook niet-radioactieve testen voor DNA-methylatie ontwikkeld. Bijvoorbeeld, assays die methylgevoelige restrictie-endonucleasen en elektroforese gebruiken om de spijsverteringsproducten te scheiden, zijn beschreven35,36. Dit soort discontinue, meerstapstests zijn niet gemakkelijk vatbaar voor medicijnontdekking. Sinds het midden van de jaren 2000 zijn verschillende DNA-methylatietests met een hogere doorvoer ontwikkeld28. Een scintillatie-nabijheidstest werd gebruikt om te screenen op DNMT1-remmers37. Een andere test met behulp van een methylgevoelige restrictie-endonuclease werd gebruikt om te screenen op DNMT3a-remmers25,38. Hoewel beide assays een hogere doorvoer mogelijk maakten dan traditionele DNA-methylatietests, vereisen de assays meerdere stappen en laten ze de observatie van methylatieactiviteit in realtime niet toe. Meer recent is een continue kinetische test beschreven die de vorming van S-adenosylhomocysteïne (SAH), een product van de methylatiereactie, koppelt aan de spectroscopische verandering bij 340 nm geassocieerd met NADPH-oxidatie39. Deze test maakt gebruik van drie koppelende enzymen om een spectroscopisch signaal te genereren.
We ontwikkelden een op fluorescentie gebaseerde endonuclease-gekoppelde DNA-methylatietest die gebruik maakt van een enkel commercieel beschikbaar koppelingsenzym en in realtime gegevens kan genereren (figuur 1). Een haarspeld oligonucleotide met drie methylcytosines wordt gebruikt als substraat. Het substraat-DNA bevat een fluorofoor aan het 5′-uiteinde en een quencher aan het 3′-uiteinde. Methylering van de gehemimethyleerde CpG-site genereert de splitsingsplaats voor de endonuclease Gla I – volledig gemethyleerde GCGC. Gla I splitsing van het product oligonucleotide geeft de fluorofoor vrij uit de quencher en genereert fluorescentie in real time. De test kan worden gebruikt om de activiteit van elke isovorm van DNMT te onderzoeken; er wordt echter een hogere activiteit waargenomen met DNMT1, omdat dit isozym bij voorkeur gehemimethyleerd DNA 1,5 methyleert. Nog robuustere activiteit wordt waargenomen als het RFTS-domein (Autoinhibitory Replication Foci Targeting Sequence) wordt verwijderd uit DNMT1. Dit domein, gevonden in het N-terminale regulatoire gebied, bindt aan de katalytische plaats en voorkomt DNA-binding. Verwijdering van de eerste ~ 600 aminozuren resulteert in een afgeknotte enzym dat aanzienlijk actiever is dan het enzym over de volledige lengte (~ 640-voudige toename van kcat / Km)40. Deze geactiveerde vorm van het enzym, aangeduid als RFTS-ontbrekende DNMT1 (aminozuren 621-1616), maakt de gemakkelijkere identificatie van remmers mogelijk vanwege de verhoogde katalytische kracht. Dit artikel presenteert een protocol om RFTS-ontbrekende DNMT1 te gebruiken in assays om te screenen op potentiële remmers van kleine moleculen. Met behulp van de endonuclease-gekoppelde continue assay wordt de beginsnelheid bepaald in de aan- en afwezigheid van enkele kleine moleculen. Elke potentiële remmer wordt onderzocht bij twee concentraties om te zoeken naar concentratieafhankelijke DNMT1-remming. Het percentage activiteit waargenomen in de aanwezigheid van de kleine moleculen werd in elk geval berekend.
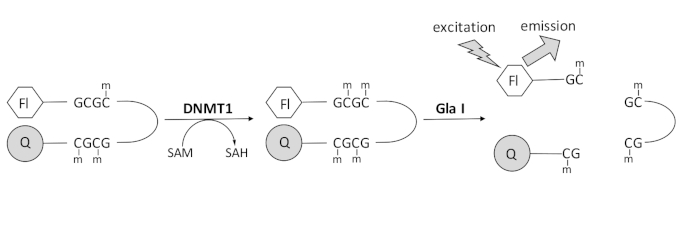
Figuur 1: DNA-methylatietest. Een gehemimethyleerd haarspeld-DNA met een fluorofoor aan het 5′-uiteinde en een quencher aan het 3′-uiteinde wordt als substraat gebruikt. DNMT1 katalyseert de overdracht van de methylgroep van S-adenosylmethionine naar de niet-gemethyleerde CpG-site, waarbij S-adenosylhomocysteïne en volledig gemethyleerd DNA wordt gegenereerd. Het DNA-product bevat de splitsingsplaats voor het endonuclease Gla I, dat volledig gemethyleerde GCGC-sites splijt. Splitsing van het product DNA geeft de 5′ fluorofoor vrij uit de 3′ quencher, waardoor fluorescentie ontstaat. Afkortingen: Fl = fluorofoor; Q =quencher; DNMT1 = DNA methyltransferase 1; SAM = S-adenosylmethionine; SAH = S-adenosylhomocysteïne. Klik hier om een grotere versie van deze figuur te bekijken.
