La méthylation de l’ADN est une marque épigénétique importante qui régule l’expression des gènes et la structure de la chromatine. La méthylation se produit principalement dans les dinucléotides CpG – cytosine suivie de guanosine; Le groupe méthyle est ajouté à la position 5 de la cytosine. Des schémas de méthylation corrects de l’ADN, et donc une expression génique appropriée, sont nécessaires pour un développement et une fonction cellulaires appropriés. De nombreux états pathologiques ont été associés à des modifications du schéma normal de méthylation 1,2,3. Par exemple, il existe un lien entre l’initiation et la progression du cancer et les altérations du schéma de méthylation de l’ADN. En règle générale, les cellules cancéreuses présentent des niveaux globaux plus faibles de méthylcytosine, ce qui contribue à l’instabilité du génome. Dans le même temps, la méthylcytosine présente dans le génome est concentrée dans les régions promotrices des gènes suppresseurs de tumeurs, ce qui conduit au silençage génique de ces protéines importantes. Notamment, les changements épigénétiques sont dynamiques et réversibles, contrairement aux mutations de l’ADN associées à la tumorigenèse. Cela a rendu les protéines impliquées dans la régulation des gènes épigénétiques intéressantescibles médicamenteuses 2,4.
Les ADN méthyltransférases (DNMT) sont les protéines responsables de la génération et du maintien des schémas de méthylation de l’ADN. Trois isoenzymes catalytiquement actifs, DNMT1, DNMT3a et DNMT3b, existent chez l’homme. Au cours du développement et de la différenciation, les méthyltransférases de novo, DNMT3a et DNMT3b, établissent des schémas de méthylation. Les deux enzymes peuvent lier la protéine DNMT3L catalytiquement inactive pour former des complexes qui présentent une activité accrue 1,5. Après la division cellulaire, les cellules filles contiennent de l’ADN hémiméthylé – de l’ADN contenant de la méthylcytosine dans un seul brin du duplex – parce que l’ADN nouvellement synthétisé est dépourvu de marques de méthylation. La fonction principale de DNMT1 est de méthyler cet ADN hémiméthylé, rétablissant ainsi le schéma de méthylation complet 1,5.
Les liens entre l’activité DNMT et le cancer sont bien établis. La surexpression de DNMT1, que ce soit par des mécanismes transcriptionnels ou post-traductionnels, est une conséquence de plusieurs voies oncogéniques courantes 6,7,8,9. Les approches génétiques visant à réduire l’activité de DNMT1 à l’aide d’allèles hypomorphiques entraînent une diminution de la formation tumorale chez les souris Apc(Min)10. Les oligonucléotides antisens qui neutralisent DNMT1 inhibent la néoplasie dans la culture cellulaire et les modèles tumorauxde souris 11,12. Ainsi, l’inhibition de l’activité DNMT1 semble être une approche prometteuse de traitement du cancer. Cependant, les rôles que jouent les isoenzymes DNMT3 ne sont pas si simples. Les mutations DNMT3a sont retrouvées dans la leucémie myéloïde aiguë13 et le syndrome myélodysplasique14. Il a été démontré qu’au moins une des mutations identifiées diminue l’activité de méthylation de l’ADN de l’enzyme15. Cependant, DNMT3b est surexprimé dans le cancer du sein16 et le cancer colorectal17. Les différents isoenzymes DNMT jouant différents rôles dans la cancérogenèse, l’identification des inhibiteurs spécifiques de l’isozyme sera essentielle. Non seulement ces composés seront utiles pour le développement de produits thérapeutiques, mais les inhibiteurs spécifiques à l’isozyme seraient également un outil précieux pour disséquer le rôle de chaque isozyme DNMT dans l’étiologie du cancer.
Plusieurs inhibiteurs de DNMT ont été rapportés dans la littérature. Les inhibiteurs connus de la DNMT peuvent être divisés en deux classes : nucléosidiques et non nucléosidiques. Les inhibiteurs nucléosidiques sont généralement des analogues de la cytidine. Ces composés sont incorporés dans l’ADN et piègent par covalence les DNMT. La 5-azacytidine et la 5-aza-2′-désoxycytidine ont été approuvées pour le traitement du syndrome myélodysplasique et de la leucémie myéloïde aiguë 4,18. La toxicité élevée, la faible biodisponibilité et l’instabilité chimique de ces composés posent des problèmes. Les travaux en cours examinent l’efficacité de la prochaine génération d’inhibiteurs nucléosidiques; SGI-110, dérivé de la 5-aza-2′-désoxycytidine, en est un exemple19,20. Les inhibiteurs nucléosidiques ne sont pas spécifiques à l’isozyme et inactiveront tout isozyme DNMT rencontré. Par conséquent, le traitement avec un agent déméthylant des nucléosides entraîne l’épuisement de tous les isoenzymes DNMT 4,18. Les inhibiteurs non nucléosidiques n’ont pas besoin d’être incorporés dans l’ADN pour exercer leurs effets inhibiteurs. Au lieu de cela, ces molécules se lient directement aux DNMT, introduisant la possibilité d’une inhibition spécifique de l’isozyme. Plusieurs inhibiteurs non nucléosidiques ont été découverts à ce jour, notamment SGI-1027 21, hydralazine22, procaïnamide 23, RG108 et dérivés 24, et des produits naturels, (−)-épigallocatéchine 3-gallate (EGCG)25 et acide laccaïque A26,27. La plupart des inhibiteurs non nucléosidiques découverts à ce jour ne sont pas isozymes-sélectifs ou présentent de faibles préférences pour un isozyme DNMT. De plus, la puissance de ces molécules doit être améliorée, en particulier dans les cellules 4,18. Il est donc nécessaire de découvrir ou de développer des inhibiteurs de DNMT isozymes-sélectifs plus puissants.
Un obstacle à la découverte de nouvelles petites molécules inhibitrices de DNMT est les tests laborieux traditionnellement utilisés pour examiner l’activité DNMT28. Les tests sont généralement discontinus avec plusieurs étapes. L’activité enzymatique des DNMT est encore systématiquement dosée à l’aide de S-adénosylméthionine (SAM) radioactive29,30,31,32,33,34. Des tests non radioactifs pour la méthylation de l’ADN ont également été développés. Par exemple, des essais utilisant des endonucléases de restriction méthylsensibles et l’électrophorèse pour séparer les produits de digestion ont été décrits35,36. Ces types de tests discontinus en plusieurs étapes ne se prêtent pas facilement à la découverte de médicaments. Depuis le milieu des années 2000, plusieurs tests de méthylation de l’ADN avec un débit plus élevé ont été développés28. Un test de proximité par scintillation a été utilisé pour dépister les inhibiteurs de DNMT137. Un autre essai utilisant une endonucléase de restriction méthylsensible a été utilisé pour dépister les inhibiteurs de DNMT3a25,38. Bien que les deux essais aient permis un débit plus élevé que les tests traditionnels de méthylation de l’ADN, les essais nécessitent plusieurs étapes et ne permettent pas d’observer l’activité de méthylation en temps réel. Plus récemment, un essai cinétique continu a été décrit qui couple la formation de S-adénosylhomocystéine (SAH), un produit de la réaction de méthylation, au changement spectroscopique à 340 nm associé à l’oxydation du NADPH39. Ce test utilise trois enzymes de couplage pour générer un signal spectroscopique.
Nous avons développé un test de méthylation de l’ADN couplé à l’endonucléase basé sur la fluorescence qui utilise une seule enzyme de couplage disponible dans le commerce et peut générer des données en temps réel (Figure 1). Un oligonucléotide en épingle à cheveux contenant trois méthylcytosines est utilisé comme substrat. L’ADN du substrat contient un fluorophore à l’extrémité 5′ et un quencher à l’extrémité 3′. La méthylation du site CpG hémiméthylé génère le site de clivage de l’endonucléase Gla I — GCGC entièrement méthylée. Le clivage Gla I de l’oligonucléotide produit libère le fluorophore de l’anti-quencher et génère une fluorescence en temps réel. Le test peut être utilisé pour examiner l’activité de toute isoforme de DNMT; cependant, une activité plus élevée est observée avec DNMT1 car cet isozyme méthyle préférentiellement l’ADN hémiméthylé 1,5. Une activité encore plus robuste est observée si le domaine autoinhibiteur de la séquence de ciblage des foyers de réplication (RFTS) est supprimé de DNMT1. Ce domaine, trouvé dans la région régulatrice N-terminale, se lie au site catalytique et empêche la liaison à l’ADN. L’élimination des premiers ~600 acides aminés donne une enzyme tronquée qui est significativement plus active que l’enzyme pleine longueur (~640 fois plus de kcat/Km)40. Cette forme activée de l’enzyme, appelée DNMT1 dépourvu de RFTS (acides aminés 621-1616), permet d’identifier plus facilement les inhibiteurs en raison de son pouvoir catalytique accru. Cet article présente un protocole d’utilisation du DNMT1 dépourvu de RFTS dans les tests de dépistage des inhibiteurs potentiels de petites molécules. À l’aide du test continu couplé à l’endonucléase, la vitesse initiale est déterminée en présence et en l’absence de quelques petites molécules. Chaque inhibiteur potentiel est examiné à deux concentrations pour rechercher une inhibition du DNMT1 dépendante de la concentration. Le pourcentage d’activité observé en présence des petites molécules a été calculé dans chaque cas.
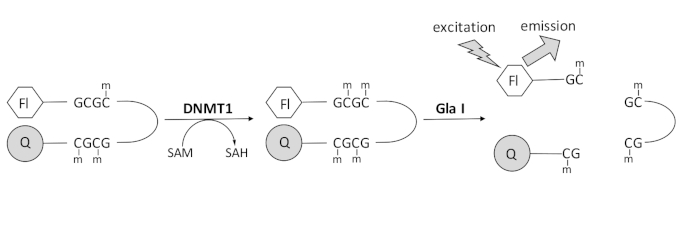
Figure 1 : Essai de méthylation de l’ADN. Un ADN d’épingle à cheveux hémiméthylé avec un fluorophore à l’extrémité 5′ et un désincant à l’extrémité 3′ est utilisé comme substrat. DNMT1 catalyse le transfert du groupe méthyle de la S-adénosylméthionine au site CpG non méthylé, générant de la S-adénosylhomocystéine et de l’ADN entièrement méthylé. Le produit d’ADN contient le site de clivage de l’endonucléase Gla I, qui clive les sites GCGC entièrement méthylés. Le clivage de l’ADN du produit libère le fluorophore 5′ de l’anti-quencher 3′, générant une fluorescence. Abréviations : Fl = fluorophore; Q = quencher; DNMT1 = ADN méthyltransférase 1; SAM = S-adénosylméthionine; HSA = S-adénosylhomocystéine. Veuillez cliquer ici pour voir une version agrandie de cette figure.
