DNAメチル化は、遺伝子発現とクロマチン構造を調節する重要なエピジェネティックマークです。メチル化は主にCpGジヌクレオチド – シトシン、続いてグアノシンで起こる。メチル基はシトシンの5位に付加される。適切な細胞の発生と機能には、正しいDNAメチル化パターン、したがって適切な遺伝子発現が必要です。多くの疾患状態は、正常なメチル化パターンの変化と関連している1,2,3。たとえば、がんの開始と進行、およびDNAメチル化パターンの変化との間には関連性があります。通常、癌細胞はメチルシトシンの全体的なレベルが低く、これがゲノム不安定性の一因となります。同時に、ゲノム中に存在するメチルシトシンは、腫瘍抑制遺伝子のプロモーター領域に集中しており、これらの重要なタンパク質の遺伝子サイレンシングをもたらす。特に、エピジェネティックな変化は、腫瘍形成に関連するDNA変異とは異なり、動的で可逆的です。これにより、エピジェネティックな遺伝子制御に関与するタンパク質が興味深い薬物標的になりました2,4。
DNAメチルトランスフェラーゼ(DNMT)は、DNAメチル化パターンの生成と維持に関与するタンパク質です。3つの触媒活性アイソザイム、DNMT1、DNMT3a、およびDNMT3bがヒトに存在します。発生および分化の過程で、de novoメチルトランスフェラーゼであるDNMT3aおよびDNMT3bはメチル化パターンを確立します。両方の酵素は、触媒的に不活性なDNMT3Lタンパク質に結合して、増加した活性を示す複合体を形成することができます1,5。細胞分裂後、娘細胞にはヘミメチル化DNA(二重鎖の1本鎖のみにメチルシトシンを含むDNA)が含まれていますが、これは新しく合成されたDNAにメチル化マークがないためです。DNMT1の主な機能は、このヘミメチル化DNAをメチル化し、完全なメチル化パターンを再確立することです1,5。
DNMT活性とがんとの関連は十分に確立されています。転写機構または翻訳後機構のいずれかによるDNMT1の過剰発現は、いくつかの一般的な発癌経路の結果である6、7、8、9。低形態対立遺伝子を用いたDNMT1活性を低下させる遺伝的アプローチは、Apc(Min)マウスにおける腫瘍形成の減少をもたらす10。DNMT1をノックダウンするアンチセンスオリゴヌクレオチドは、細胞培養およびマウス腫瘍モデルにおける新生物を阻害する11、12。したがって、DNMT1活性を阻害することは、有望な癌治療アプローチのように思われます。しかし、DNMT3アイソザイムが果たす役割はそれほど単純ではありません。DNMT3a変異は、急性骨髄性白血病13および骨髄異形成症候群14に見られます。同定された変異の少なくとも1つは、酵素15のDNAメチル化活性を低下させることが示されている。しかし、DNMT3bは乳癌16および結腸直腸癌17において過剰発現している。さまざまなDNMTアイソザイムが発がんにおいて異なる役割を果たしているため、アイソザイム特異的阻害剤の同定が重要になります。これらの化合物は治療薬の開発に役立つだけでなく、アイソザイム特異的阻害剤は、がんの病因における各DNMTアイソザイムの役割を分析するための非常に貴重なツールにもなります。
いくつかのDNMT阻害剤が文献で報告されています。既知のDNMT阻害剤は、ヌクレオシドと非ヌクレオシドの2つのクラスに分けることができます。ヌクレオシド阻害剤は、典型的にはシチジン類似体である。これらの化合物はDNAに組み込まれ、DNMTを共有結合的にトラップします。 5-アザシチジンおよび5-アザ-2′-デオキシシチジンは、骨髄異形成症候群および急性骨髄性白血病の治療薬として承認されています4,18。これらの化合物の高い毒性、低いバイオアベイラビリティ、および化学的不安定性は問題を提示します。進行中の研究は、次世代のヌクレオシド阻害剤の有効性を調べることです。SGI−110は、5−アザ−2’−デオキシシチジンから誘導される、19、20の一例である。ヌクレオシド阻害剤はアイソザイム特異的ではなく、遭遇したDNMTアイソザイムを不活性化します。したがって、ヌクレオシド脱メチル化剤で処理すると、すべてのDNMTアイソザイムが枯渇します4,18。非ヌクレオシド阻害剤は、その阻害効果を発揮するためにDNAに組み込む必要はありません。代わりに、これらの分子はDNMTに直接結合し、アイソザイム特異的阻害の可能性をもたらします。SGI-1027 21、ヒドララジン22、プロカインアミド23、RG108および誘導体24、ならびに天然物、(−)−エピガロカテキン3−ガレート(EGCG)25およびラッカイン酸A26,27を含む、いくつかの非ヌクレオシド阻害剤がこれまでに発見されている。これまでに発見された非ヌクレオシド阻害剤のほとんどは、アイソザイム選択的ではないか、1つのDNMTアイソザイムに対して弱い選好を示します。さらに、これらの分子の効力は、特に細胞4,18において改善される必要がある。したがって、より強力なアイソザイム選択的DNMT阻害剤を発見または開発する必要があります。
DNMTの新しい低分子阻害剤を発見するためのハードルは、DNMT活性を調べるために伝統的に使用されている骨の折れるアッセイです28。アッセイは通常、複数のステップで不連続です。DNMTの酵素活性は、放射性S-アデノシルメチオニン(SAM)29,30,31,32,33,34を用いて日常的にアッセイされています。DNAメチル化のための非放射性アッセイも開発されています。例えば、消化産物を分離するためにメチル感受性制限エンドヌクレアーゼおよび電気泳動を利用するアッセイが記載されている35、36。これらのタイプの不連続な多段階アッセイは、創薬に容易に適していません。2000年代半ば以降、より高いスループットを有するいくつかのDNAメチル化アッセイが開発されてきた28。シンチレーション近接アッセイを用いて、DNMT1阻害剤37をスクリーニングした。メチル感受性制限エンドヌクレアーゼを利用する別のアッセイを、DNMT3a阻害剤のスクリーニングに使用した25、38。どちらのアッセイも従来のDNAメチル化アッセイよりも高いスループットを可能にしましたが、アッセイは複数のステップを必要とし、メチル化活性をリアルタイムで観察することはできません。最近では、メチル化反応の1つの生成物であるS-アデノシルホモシステイン(SAH)の形成を、NADPH酸化に関連する340 nmでの分光学的変化に結合する連続速度論アッセイが記載されている39。このアッセイでは、3つのカップリング酵素を利用して分光シグナルを生成します。
我々は、市販の単一のカップリング酵素を利用し、リアルタイムでデータを生成できる蛍光ベースのエンドヌクレアーゼ共役DNAメチル化アッセイを開発しました(図1)。3つのメチルシトシンを含むヘアピンオリゴヌクレオチドを基質として用いる。基質DNAには、5’末端に蛍光色素、3’末端に消光物質が含まれています。ヘミメチル化CpGサイトのメチル化は、エンドヌクレアーゼGla I – 完全メチル化GCGCの切断部位を生成する。生成物オリゴヌクレオチドのGla I切断は、クエンチャーから蛍光色素を放出し、リアルタイムで蛍光を生成します。このアッセイは、DNMTの任意のアイソフォームの活性を調べるために使用できます。しかし、このアイソザイムがヘミメチル化DNA1,5を優先的にメチル化するため、DNMT1ではより高い活性が観察されます。自己抑制性複製病巣標的配列(RFTS)ドメインがDNMT1から除去されると、さらに堅牢な活性が観察されます。N末端調節領域に見られるこのドメインは、触媒部位に結合し、DNA結合を妨げます。最初の~600個のアミノ酸を除去すると、完全長酵素よりも有意に活性の高い切断酵素が得られます(kcat/Kmの~640倍の増加)40。RFTS欠損DNMT1(アミノ酸621〜1616)と呼ばれるこの活性化型の酵素は、触媒力の増加により、阻害剤の同定を容易にします。この論文は、潜在的な低分子阻害剤をスクリーニングするためのアッセイでRFTS欠損DNMT1を利用するためのプロトコルを提示します。エンドヌクレアーゼ共役連続アッセイを用いて、数個の低分子の存在下および非存在下で初速度を決定する。各潜在的な阻害剤を2つの濃度で調べて、濃度依存的なDNMT1阻害を探します。小分子の存在下で観察された活性パーセントを、それぞれの場合において計算した。
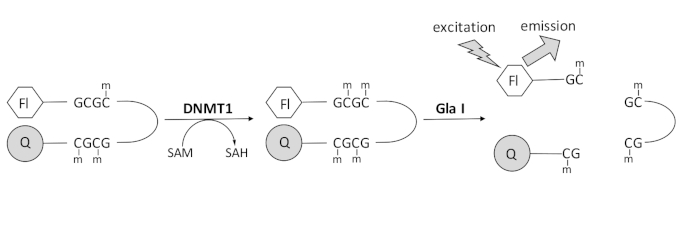
図1:DNAメチル化アッセイ。 5’末端に蛍光色素、3’末端に消光物質を有するヘミメチル化ヘアピンDNAを基質として使用します。DNMT1は、 S-アデノシルメチオニンから非メチル化CpG部位へのメチル基の転移を触媒し、 S-アデノシルホモシステインおよび完全メチル化DNAを生成します。DNA産物には、完全にメチル化されたGCGC部位を切断するエンドヌクレアーゼGla Iの切断部位が含まれています。製品DNAが切断されると、3’クエンチャーから5’蛍光色素が放出され、蛍光が発生します。略語:Fl =フルオロフォア;Q =クエンチャー;DNMT1 = DNAメチルトランスフェラーゼ1;SAM = S-アデノシルメチオニン;SAH = S-アデノシルホモシステイン。 この図の拡大版を表示するには、ここをクリックしてください。
