DNA-metylering är ett viktigt epigenetiskt märke som reglerar genuttryck och kromatinstruktur. Metylering sker huvudsakligen i CpG-dinukleotider – cytosin följt av guanosin; Metylgruppen tillsätts till 5-positionen av cytosin. Korrekta DNA-metyleringsmönster, och därmed korrekt genuttryck, behövs för lämplig cellulär utveckling och funktion. Många sjukdomstillstånd har associerats med förändringar i det normala metyleringsmönstret 1,2,3. Till exempel finns det en koppling mellan cancerinitiering och progression och förändringar i DNA-metyleringsmönstret. Vanligtvis uppvisar cancerceller lägre totala nivåer av metylcytosin, vilket bidrar till genominstabilitet. Samtidigt koncentreras metylcytosinet som är närvarande i genomet i promotorregionerna av tumörsuppressorgener, vilket leder till gendämpning av dessa viktiga proteiner. I synnerhet är epigenetiska förändringar dynamiska och reversibla, till skillnad från DNA-mutationerna associerade med tumorigenes. Detta har gjort proteinerna som är involverade i epigenetisk genreglering intressanta läkemedelsmål 2,4.
DNA-metyltransferaser (DNMT) är de proteiner som är ansvariga för att generera och upprätthålla DNA-metyleringsmönster. Tre katalytiskt aktiva isozymer, DNMT1, DNMT3a och DNMT3b, finns hos människor. Under utveckling och differentiering etablerar de novo-metyltransferaserna, DNMT3a och DNMT3b, metyleringsmönster. Båda enzymerna kan binda det katalytiskt inaktiva DNMT3L-proteinet för att bilda komplex som uppvisar ökad aktivitet 1,5. Efter celldelning innehåller dotterceller hemimetylerat DNA – DNA som innehåller metylcytosin i endast en sträng av duplexen – eftersom det nyligen syntetiserade DNA saknar metyleringsmärken. Den huvudsakliga funktionen hos DNMT1 är att metylera detta hemimetylerade DNA och därmed återupprätta det fullständiga metyleringsmönstret 1,5.
Kopplingar mellan DNMT-aktivitet och cancer är väl etablerade. Överuttryck av DNMT1, antingen genom transkriptionella eller post-translationella mekanismer, är en följd av flera vanliga onkogena vägar 6,7,8,9. Genetiska tillvägagångssätt för att sänka DNMT1-aktivitet med hypomorfa alleler resulterar i minskad tumörbildning hos Apc (Min) möss10. Antisense oligonukleotider som slår ner DNMT1 hämmar neoplasi i cellodling och mustumörmodeller11,12. Således verkar hämmande DNMT1-aktivitet som en lovande cancerterapimetod. De roller som DNMT3-isozymerna spelar är dock inte så enkla. DNMT3a-mutationer finns i akut myeloisk leukemi13 och myelodysplastiskt syndrom14. Minst en av de identifierade mutationerna har visat sig minska DNA-metyleringsaktiviteten hos enzymet15. DNMT3b är dock överuttryckt i bröstcancer16 och kolorektal cancer17. Eftersom de olika DNMT-isozymerna spelar olika roller i cancerframkallande kommer det att vara avgörande att identifiera isozymspecifika hämmare. Dessa föreningar kommer inte bara att vara användbara för utveckling av terapier, utan isozymspecifika hämmare skulle också vara ett ovärderligt verktyg för att dissekera rollen för varje DNMT-isozym i canceretiologi.
Flera DNMT-hämmare har rapporterats i litteraturen. Kända DNMT-hämmare kan delas in i två klasser: nukleosid och icke-nukleosid. Nukleosidhämmare är vanligtvis cytidinanaloger. Dessa föreningar införlivas i DNA och kovalent fälla DNMTs. 5-azacytidin och 5-aza-2′-deoxycytidin har godkänts för behandling av myelodysplastiskt syndrom och akut myeloisk leukemi 4,18. Den höga toxiciteten, låg biotillgänglighet, och kemisk instabilitet av dessa föreningar ger problem. Pågående arbete undersöker effekten av nästa generations nukleosidhämmare; SGI-110, härledd från 5-aza-2′-deoxicytidin, är ett exempel19,20. Nukleosidhämmare är inte isozymspecifika och kommer att inaktivera alla DNMT-isozym som påträffas. Därför resulterar behandling med ett nukleosiddemetyleringsmedel i uttömning av alla DNMT-isozymer 4,18. Icke-nukleosidhämmare behöver inte införlivas i DNA för att utöva sina hämmande effekter. Istället binder dessa molekyler direkt till DNMT, vilket introducerar möjligheten till isozymspecifik hämning. Flera icke-nukleosidhämmare har hittills upptäckts, inklusive SGI-1027 21, hydralazin 22, prokainamid23, RG108 och derivat 24, och naturliga produkter, (−)-epigallocatechin 3-gallat (EGCG)25 och lakcainsyra A 26,27. De flesta av de icke-nukleosidhämmare som hittills upptäckts är inte isozymeselektiva eller visar svaga preferenser för ett DNMT-isozym. Dessutom måste styrkan hos dessa molekyler förbättras, särskilt i cellerna 4,18. Således finns det ett behov av att upptäcka eller utveckla mer potenta, isozymselektiva DNMT-hämmare.
Ett hinder för att upptäcka nya småmolekylära hämmare av DNMT är de mödosamma analyser som traditionellt används för att undersöka DNMT-aktivitet28. Analyser är vanligtvis diskontinuerliga med flera steg. Den enzymatiska aktiviteten hos DNMT analyseras fortfarande rutinmässigt med radioaktivt S-adenosylmetionin (SAM)29,30,31,32,33,34. Icke-radioaktiva analyser för DNA-metylering har också utvecklats. Till exempel har analyser som använder metylkänsliga restriktionsendonukleaser och elektrofores för att separera matsmältningsprodukterna beskrivits35,36. Dessa typer av diskontinuerliga, flerstegsanalyser är inte lätt mottagliga för läkemedelsupptäckt. Sedan mitten av 2000-talet har flera DNA-metyleringsanalyser med högre genomströmning utvecklats28. En scintillationsnärhetsanalys användes för att screena för DNMT1-hämmare37. En annan analys som använder ett metylkänsligt restriktionsendonukleas användes för att screena för DNMT3a-hämmare25,38. Medan båda analyserna möjliggjorde högre genomströmning än traditionella DNA-metyleringsanalyser, kräver analyserna flera steg och tillåter inte observation av metyleringsaktivitet i realtid. På senare tid har en kontinuerlig kinetikanalys beskrivits som kopplar bildningen av S-adenosylhomocystein (SAH), en produkt av metyleringsreaktionen, till den spektroskopiska förändringen vid 340 nm associerad med NADPH-oxidation39. Denna analys använder tre kopplingsenzymer för att generera en spektroskopisk signal.
Vi utvecklade en fluorescensbaserad endonukleaskopplad DNA-metyleringsanalys som använder ett enda kommersiellt tillgängligt kopplingsenzym och kan generera data i realtid (figur 1). En hårnåloligonukleotid innehållande tre metylcytosiner används som substrat. Substrat-DNA innehåller en fluorofor på 5′-änden och en släckare på 3′-änden. Metylering av det hemimetylerade CpG-stället genererar klyvningsstället för endonukleas Gla I – helt metylerad GCGC. Gla I-klyvning av produkten oligonukleotid frigör fluoroforen från släckaren och genererar fluorescens i realtid. Analysen kan användas för att undersöka aktiviteten hos vilken isoform av DNMT som helst; emellertid observeras högre aktivitet med DNMT1 eftersom detta isozym företrädesvis metylerar hemimetylerat DNA 1,5. Ännu mer robust aktivitet observeras om den autoinhibitoriska RFTS-domänen (Replication Foci Targeting Sequence) tas bort från DNMT1. Denna domän, som finns i N-terminalregleringsregionen, binder till den katalytiska platsen och förhindrar DNA-bindning. Avlägsnande av de första ~ 600 aminosyrorna resulterar i ett stympat enzym som är signifikant mer aktivt än fullängdsenzymet (~ 640-faldig ökning av kkatt / Km) 40. Denna aktiverade form av enzymet, kallad RFTS-saknad DNMT1 (aminosyror 621–1616), möjliggör enklare identifiering av hämmare på grund av dess ökade katalytiska kraft. Detta dokument presenterar ett protokoll för att använda RFTS-saknade DNMT1 i analyser för att screena för potentiella småmolekylära hämmare. Med hjälp av den endonukleaskopplade kontinuerliga analysen bestäms initialhastigheten i närvaro och frånvaro av några små molekyler. Varje potentiell hämmare undersöks vid två koncentrationer för att leta efter koncentrationsberoende DNMT1-hämning. Den procentuella aktiviteten som observerades i närvaro av de små molekylerna beräknades i varje enskilt fall.
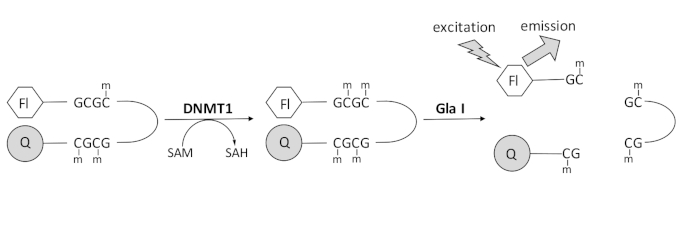
Figur 1: DNA-metyleringsanalys. Ett hemimetylerat hårnåls-DNA med en fluorofor i 5′-änden och en släckare på 3′-änden används som substrat. DNMT1 katalyserar överföringen av metylgruppen från S-adenosylmetionin till det icke-metylerade CpG-stället, vilket genererar S-adenosylhomocystein och fullständigt metylerat DNA. DNA-produkten innehåller klyvningsstället för endonukleas Gla I, som klyver helt metylerade GCGC-platser. Klyvning av produkten DNA frigör 5′-fluoroforen från 3′-släckaren och genererar fluorescens. Förkortningar: Fl = fluorofor; Q =släckare; DNMT1 = DNA-metyltransferas 1; SAM = S-adenosylmetionin; SAH = S-adenosylhomocystein. Klicka här för att se en större version av denna siffra.
