Биомолекулярные взаимодействия:
Белки являются неотъемлемой частью организмов и участвуют в многочисленных молекулярных путях, таких как клеточный метаболизм, клеточная структура, клеточная сигнализация, иммунные реакции, клеточная адгезия и многое другое. В то время как некоторые белки выполняют свою функцию (функции) независимо, большинство белков взаимодействуют с другими белками, используя связующий интерфейс для координации надлежащей биологической активности1.
Биомолекулярные взаимодействия могут быть в основном классифицированы на основе различных структурных и функциональных характеристик вовлеченных белков2, например, на основе белковых поверхностей, комплексной стабильности или персистенции взаимодействий3. Идентификация основных белков и их роли в биомолекулярных взаимодействиях имеет жизненно важное значение для понимания биохимических механизмов на молекулярном уровне4. В настоящее время существуют различные подходы к обнаружению этих взаимодействий5: in vitro6, in silico7, в живых клетках8, ex vivo9 и in vivo10, причем каждый из них имеет свои сильные и слабые стороны.
Анализы in vivo выполняются с использованием всего животного в качестве экспериментального инструмента11, а анализыt he ex vivo выполняются на экстрактах тканей или целых органов (например, сердце, мозг, печень) в контролируемой внешней среде, обеспечивая минимальные изменения в естественных условиях. Наиболее распространенным применением исследований in vivo и ex vivo является оценка фармакокинетики, фармакодинамики и токсичности потенциальных фармакологических агентов до испытаний на людях путем обеспечения их общей безопасности и эффективности12.
Биомолекулярные взаимодействия также могут быть обнаружены в живых клетках. Визуализация живых клеток позволяет нам наблюдать динамические взаимодействия, поскольку они выполняют реакции определенного биохимического пути13. Кроме того, методы обнаружения, такие как биолюминесценция или флуоресцентный резонансный перенос энергии, могут предоставить информацию о том, где и когда эти взаимодействия происходят в клетке14. Хотя обнаружение в живых клетках предлагает важные детали, эти методологии обнаружения основаны на оптике и метках, которые могут не отражать нативную биологию; они также менее контролируемы, чем методы in vitro , и требуют специализированных знаний для выполнения15.
Вычислительные методы in silico в основном используются для крупномасштабного скрининга молекул-мишеней перед экспериментами in vitro . Вычислительные методы прогнозирования, компьютерные базы данных, молекулярная стыковка, количественные отношения структура-активность и другие подходы к моделированию молекулярной динамики являются одними из хорошо зарекомендовавших себя инструментов in silico 16. По сравнению с трудоемкими экспериментальными методами, инструменты in silico могут легко делать прогнозы с высокой чувствительностью, но с пониженной точностью в прогностической производительности17.
Анализы in vitro проводятся с микроорганизмами или биологическими молекулами вне их стандартного биологического контекста. Изображение биомолекулярных взаимодействий с помощью методов in vitro имеет решающее значение для понимания функций белка и биологии, стоящей за сложной сетью функционирования клеток. Предпочтительная методология анализа выбирается в соответствии с внутренними свойствами белка, кинетическими значениями, а также режимом и интенсивностью взаимодействий18,19.
Взаимодействие Hsp90/Cdc37:
Путь шаперон-киназы, соединяющий Hsp90 и Cdc37, является перспективной терапевтической мишенью в биологии опухоли20. Hsp90 играет центральную роль в контроле клеточного цикла, сборке белка, выживании клеток и сигнальных путях. Белки, которые полагаются на Hsp90 для своих функций, доставляются в Hsp90 для комплексообразования через ко-шаперон, такой как Cdc37. Комплекс Hsp90/Cdc37 контролирует сворачивание большинства протеинкиназ и служит концентратором для множества внутриклеточных сигнальных сетей21. Это многообещающая противоопухолевая мишень из-за ее повышенной экспрессии при различных злокачественных новообразованиях, включая острый миелобластный лейкоз, множественную миелому и гепатоцеллюлярную карциному22,23.
Широко используемые методы обнаружения биомолекулярного взаимодействия in vitro
Коиммунопреципитация (co-IP) представляет собой метод, основанный на специфичности антиген-антитело для идентификации биологически значимых взаимодействий24. Основным недостатком этого метода является его неспособность обнаруживать низкоаффинные взаимодействия и кинетические значения24. Биофизические методы, такие как изотермическая титрующая калориметрия (ITC), поверхностный плазмонный резонанс (SPR), биослойная интерферометрия (BLI) и технология FEB, являются предпочтительными для определения кинетических значений.
ITC представляет собой биофизический метод обнаружения, основанный на определении энергии связывания наряду с полным термодинамическим анализом для характеристики биомолекулярных взаимодействий25. Основным преимуществом ITC является то, что он не требует какой-либо маркировки или фиксации целевого белка. Основными трудностями, с которыми сталкивается ITC, являются высокая концентрация целевого белка, необходимого для одного эксперимента, и сложность анализа нековалентных комплексов из-за небольших энтальпийсвязывания 26. Как SPR, так и BLI являются биофизическими методами без меток, которые полагаются на иммобилизацию молекулы-мишени на поверхности датчика с последующими инъекциями аналита через иммобилизованную мишень27,28. В SPR измеряются изменения показателя преломления при биомолекулярных взаимодействиях27; в BLI интерференция в отраженном свете регистрируется в режиме реального времени как изменение длины волны в зависимости от времени28. Как SPR, так и BLI имеют общие преимущества, заключающиеся в высокой специфичности, чувствительности и возможностях обнаружения29. В обоих способах белок-мишень иммобилизуется на биосенсорных поверхностях, и, следовательно, может произойти некоторая потеря нативной конформации мишени, что затрудняет различие между конкретными и неспецифическими взаимодействиями30. BLI использует дорогостоящие одноразовые волоконно-оптические биосенсоры для иммобилизации мишени и, следовательно, является дорогостоящим методом31. По сравнению с этими хорошо зарекомендовавшими себя инструментами биомолекулярного обнаружения, технология FEB предлагает надежную и свободную от этикеток платформу, используя низкие наномолярные концентрации для биомолекулярного обнаружения в режиме реального времени с кинетической характеристикой. Технология FEB также преодолевает проблемы, с которыми сталкивается ITC, и является более экономически эффективной по сравнению с SPR или BLI.
Биосенсоры на основе полевых транзисторов (FET) являются новой областью для обнаружения биомолекулярных взаимодействий, предлагая различные биомедицинские приложения. В системе FET мишени иммобилизуются в микросхемы биосенсора, а взаимодействия обнаруживаются изменениями проводимости32. Уникальной особенностью, которую следует учитывать при разработке эффективного электронного биосенсора, являются физико-химические свойства, такие как полупроводниковая природа и химическая стабильность материала покрытия, используемого для изготовления поверхностидатчика 33. Обычные материалы, такие как кремний, используемые для FET, ограничивают чувствительность датчиков, поскольку для правильного функционирования требуются оксидные слои, зажатые между транзисторным каналом и определенной средой34. Кроме того, кремниевые транзисторы чувствительны к средам с высоким содержанием соли, что затрудняет измерение биологических взаимодействий в их естественной среде. Биосенсор на основе графена представлен в качестве альтернативы, поскольку он обеспечивает отличную химическую стабильность и электрическое поле. Поскольку графен представляет собой единый атомный слой углерода, он чрезвычайно чувствителен как полупроводник и химически совместим с биологическими растворами; оба эти качества желательны для генерации совместимых электронных биосенсоров35. Замечательный сверхвысокий нагрузочный потенциал биомолекул, предлагаемых биосенсорами с графеновым покрытием, приводит к разработке технологии биосенсоров FEB на основе графена.
Принцип технологии FEB: FEB – это метод биомолекулярного обнаружения без меток, который измеряет электрический ток через графеновый биосенсор, к которому иммобилизуются связывающие мишени. Взаимодействия между иммобилизованным белком и анализируемым веществом приводят к изменениям тока, которые контролируются в режиме реального времени, что позволяет проводить точные кинетические измерения36.
Контрольно-измерительные приборы: Система FEB содержит графеновый полевой транзистор (gFET) сенсорный чип и электронный считыватель, который подает постоянное напряжение на протяжении всего эксперимента (рисунок 1). Аналит наносят в растворе на целевой белок, иммобилизованный на поверхности биосенсора. Когда происходит взаимодействие, изменение тока измеряется и записывается в режиме реального времени. По мере увеличения концентрации анализируемого вещества доля связанного аналита также будет увеличиваться, вызывая более высокие чередования в токе. С помощью автоматизированного аналитического программного обеспечения, поставляемого с прибором (Таблица материалов), I-Response измеряется и регистрируется в единицах биозондирования (BU)37. I-Response определяется как изменение тока (I) через чип биосенсора, измеренное в режиме реального времени при взаимодействии иммобилизованной мишени с анализируемым веществом. Программное обеспечение для автоматизированного анализа FEB может анализировать как I-Response, так и C-Response на события динамического взаимодействия, где C-Response регистрирует изменения емкости (C). Вариации как i-response, так и C-Response непосредственно соответствуют доле связанного аналита и могут быть дополнительно проанализированы для получения значений KD . Программное обеспечение для автоматизированного анализа по умолчанию предпочитает I-Response.
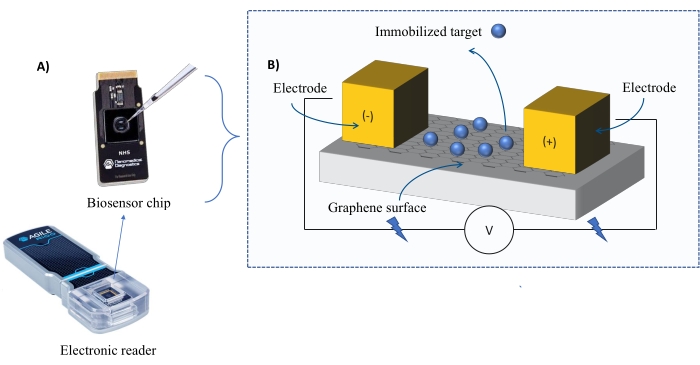
Рисунок 1: Обзор экспериментальной установки. (A) Чип на основе графена и электронный считыватель. (B) Обзор компонентов микросхемы. Чип прикреплен к двум электродам, которые подают ток в систему. Поверхность чипа покрыта графеном, который при активации может связывать мишень. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Методология:
Первоначально активированный микросхема биосенсора вставляется в устройство FEB (рисунок 1) с последующим выполнением шагов, описанных ниже: (1) Калибровка: Эксперимент начинается с калибровки системы с использованием 1x фосфатно-буферного физиологического раствора (PBS; pH = 7,4) для создания базового ответа равновесия. (2) Ассоциация: Анализируемый элемент вводится в чип, и I-Response контролируется до тех пор, пока не будет достигнуто насыщение связывания. (3) Диссоциация: анализируемый материал диссоциируется с использованием 1x PBS. (4) Регенерация: Остатки анализируемого вещества удаляются с использованием 1x PBS. (5) Стирка: В общей сложности пять стирок выполняются с использованием 1x PBS для тщательного удаления связанных и несвязанных аналитов из чипа.
Анализ:
Анализ данных выполняется с использованием полностью автоматизированного программного обеспечения, поставляемого с прибором. Автоматизированное аналитическое программное обеспечение генерирует график Hill fit со значением KD . График соответствия Хилла описывает связь анализируемого вещества с целевым белком в зависимости от концентраций анализируемого вещества. Концентрация, при которой достигается полумаксимальный отклик, пропорциональна значению KD . Низкое значение KD представляет собой высокое сродство связывания и наоборот.
Для проверки данных, полученных в ходе эксперимента FEB, I-Ответы извлекаются из каждой точки считывания для каждой концентрации анализируемого вещества с использованием программного обеспечения для анализа/экспорта данных и могут быть экспортированы в другое программное обеспечение для статистического анализа (см. Таблицу материалов), как описано ниже.
