Biomoleculaire interacties:
Eiwitten zijn essentiële onderdelen van organismen en nemen deel aan tal van moleculaire routes zoals celmetabolisme, celstructuur, celsignalering, immuunresponsen, celadhesie en meer. Hoewel sommige eiwitten hun functie(s) onafhankelijk uitvoeren, interageren de meeste eiwitten met andere eiwitten met behulp van een bindende interface om de juiste biologische activiteit te coördineren1.
Biomoleculaire interacties kunnen voornamelijk worden geclassificeerd op basis van de verschillende structurele en functionele kenmerken van betrokken eiwitten2, bijvoorbeeld op basis van de eiwitoppervlakken, de complexe stabiliteit of de persistentie van interacties3. Het identificeren van essentiële eiwitten en hun rol in biomoleculaire interacties is van vitaal belang voor het begrijpen van biochemische mechanismen op moleculair niveau4. Momenteel zijn er verschillende benaderingen om deze interacties5 te detecteren: in vitro6, in silico7, in levende cellen8, ex vivo9 en in vivo10 met elk zijn eigen sterke en zwakke punten.
De in vivo assays worden uitgevoerd met behulp van het hele dier als een experimenteel hulpmiddel11, ende ex vivo assays worden uitgevoerd op weefselextracten of hele organen (bijv. Hart, hersenen, lever) in een gecontroleerde externe omgeving door minimale veranderingen in natuurlijke omstandigheden te bieden. De meest voorkomende toepassing van in vivo en ex vivo studies is het evalueren van de farmacokinetiek, farmacodynamiek en toxiciteitseffecten van potentiële farmacologische agentia vóór proeven bij mensen door hun algehele veiligheid en werkzaamheid te waarborgen12.
Biomoleculaire interacties kunnen ook worden gedetecteerd in levende cellen. Beeldvorming van levende cellen stelt ons in staat om dynamische interacties te observeren terwijl ze de reacties van een bepaalde biochemische route uitvoeren13. Bovendien kunnen detectietechnieken, zoals bioluminescentie of fluorescentieresonantie-energieoverdracht, informatie geven over waar en wanneer deze interacties plaatsvinden in de cel14. Hoewel detectie in levende cellen cruciale details biedt, vertrouwen deze detectiemethoden op optica en labels, die mogelijk niet de inheemse biologie weerspiegelen; ze zijn ook minder gecontroleerd dan in vitro methoden en vereisen gespecialiseerde expertise om15 uit te voeren.
De in silico computationele methoden worden voornamelijk gebruikt voor grootschalige screening van doelmoleculen vóór de in vitro experimenten. Computationele voorspellingsmethoden, computergebaseerde databases, moleculaire docking, kwantitatieve structuur-activiteitsrelaties en andere moleculaire dynamica-simulatiebenaderingen behoren tot de gevestigde in silico-tools 16. In vergelijking met moeizame experimentele technieken kunnen de in silico-tools gemakkelijk voorspellingen doen met een hoge gevoeligheid, maar met verminderde nauwkeurigheid in voorspellende prestaties17.
In vitro assays worden uitgevoerd met micro-organismen of biologische moleculen buiten hun standaard biologische context. Het weergeven van biomoleculaire interacties door middel van in vitro methoden is van cruciaal belang voor het begrijpen van eiwitfuncties en de biologie achter het complexe netwerk van celfuncties. De voorkeurstestmethodologie wordt gekozen op basis van de intrinsieke eigenschappen van het eiwit, kinetische waarden en de wijze en intensiteit van interacties18,19.
De interactie Hsp90/Cdc37:
De chaperonne-kinaseroute, die Hsp90 en Cdc37 verbindt, is een veelbelovend therapeutisch doelwit in de tumorbiologie20. Hsp90 speelt een centrale rol in de controle van de celcyclus, eiwitassemblage, celoverleving en signaalroutes. Eiwitten die afhankelijk zijn van Hsp90 voor hun functies worden geleverd aan Hsp90 voor complexatie via een co-chaperonne, zoals Cdc37. Het Hsp90/Cdc37-complex regelt de vouwing van de meeste eiwitkinasen en dient als hub voor een groot aantal intracellulaire signaleringsnetwerken21. Het is een veelbelovend antitumordoel vanwege de verhoogde expressie in verschillende maligniteiten, waaronder acute myeloblastische leukemie, multipel myeloom en hepatocellulair carcinoom22,23.
Veelgebruikte in vitro biomoleculaire interactie detectietechnieken
Co-immunoprecipitatie (co-IP) is een techniek die vertrouwt op antigeen-antilichaamspecificiteit om biologisch relevante interacties te identificeren24. Het belangrijkste nadeel van deze methode is het onvermogen om interacties met lage affiniteit en kinetische waarden te detecteren24. Biofysische methoden zoals isotherme titratiecalorimetrie (ITC), oppervlakteplasmonresonantie (SPR), biolaaginterferometrie (BLI) en FEB-technologie hebben de voorkeur om de kinetische waarden te bepalen.
ITC is een biofysische detectiemethode gebaseerd op de bepaling van bindingsenergie samen met een volledige thermodynamica-analyse om biomoleculaire interacties te karakteriseren25. Het belangrijkste voordeel van ITC is dat het geen etikettering of fixatie van het doeleiwit vereist. De belangrijkste problemen die ITC ondervindt, zijn de hoge concentratie doeleiwit die nodig is voor één experiment en de moeilijkheid om niet-covalente complexen te analyseren als gevolg van kleine bindende enthalpies26. Zowel SPR als BLI zijn labelvrije biofysische technieken die afhankelijk zijn van de immobilisatie van het doelmolecuul op het sensoroppervlak, gevolgd door daaropvolgende injecties van de analyt over het geïmmobiliseerde doel27,28. In SPR worden veranderingen in de brekingsindex tijdens biomoleculaire interacties gemeten27; in BLI wordt de interferentie in gereflecteerd licht in real-time geregistreerd als een verandering in golflengte als functie van tijd28. Zowel SPR als BLI delen gemeenschappelijke voordelen van het aanbieden van hoge specificiteit, gevoeligheid en detectiemogelijkheden29. In beide methoden wordt het doeleiwit geïmmobiliseerd op biosensoroppervlakken en daarom kan er enig verlies zijn van de oorspronkelijke conformatie van het doelwit, waardoor het moeilijk is om onderscheid te maken tussen specifieke versus niet-specifieke interacties30. BLI gebruikt dure wegwerpbare glasvezelbiosensoren om het doelwit te immobiliseren en is daarom een kostbare techniek31. In vergelijking met deze gevestigde biomoleculaire detectietools biedt FEB-technologie een betrouwbaar en labelvrij platform door lage nanomolelarconcentraties te gebruiken voor biomoleculaire detectie in realtime met kinetische karakterisering. De FEB-technologie overwint ook de borrelende uitdagingen in ITC en is kosteneffectiever in vergelijking met SPR of BLI.
De op veldeffecttransistor (FET) gebaseerde biosensoren is een opkomend veld voor het detecteren van biomoleculaire interacties door gevarieerde biomedische toepassingen aan te bieden. In het FET-systeem worden doelen geïmmobiliseerd naar de biosensorchips en interacties worden gedetecteerd door veranderingen in geleiding32. Het unieke kenmerk waarmee rekening moet worden gehouden bij de ontwikkeling van een efficiënte elektronische biosensor zijn de fysisch-chemische eigenschappen zoals de semi-geleidende aard en chemische stabiliteit van het coatingmateriaal dat wordt gebruikt om het sensoroppervlak te fabriceren33. Conventionele materialen zoals silicium die voor FET worden gebruikt, hebben de gevoeligheid van sensoren beperkt omdat het oxidelagen vereist die ingeklemd zitten tussen het transistorkanaal en een specifieke omgeving voor een goede werking34. Bovendien zijn siliciumtransistors gevoelig voor zoutrijke omgevingen, waardoor het moeilijk is om biologische interacties in hun natuurlijke omgeving te meten. De op grafeen gebaseerde biosensor wordt gepresenteerd als een alternatief omdat het een uitstekende chemische stabiliteit en elektrisch veld biedt. Aangezien grafeen een enkele atomaire laag koolstof is, is het zowel extreem gevoelig als een halfgeleider en chemisch compatibel met biologische oplossingen; beide kwaliteiten zijn wenselijk om compatibele elektronische biosensoren te genereren35. Het opmerkelijke ultrahoge laadpotentieel van biomoleculen dat wordt geboden door biosensoren met grafeencoating, leidt tot de ontwikkeling van op grafeen gebaseerde biosensoren FEB-technologie.
Principe van FEB-technologie: FEB is een labelvrije biomoleculaire detectietechniek die de elektrische stroom meet door de grafeenbiosensor waaraan de bindingsdoelen worden geïmmobiliseerd. Interacties tussen het geïmmobiliseerde eiwit en de analyt resulteren in veranderingen in de stroom die in realtime worden gemonitord, waardoor nauwkeurige kinetische metingenmogelijk zijn 36.
Instrumentatie: Het FEB-systeem bestaat uit een grafeenveldeffecttransistor (gFET) sensorchip en een elektronische lezer die gedurende het hele experiment een constante spanning toepast (figuur 1). De analyt wordt in oplossing aangebracht op het doeleiwit dat op het biosensoroppervlak is geïmmobiliseerd. Wanneer een interactie optreedt, wordt een verandering in de stroom gemeten en in realtime geregistreerd. Naarmate de analytconcentratie toeneemt, zal ook de fractie van gebonden analyt toenemen, waardoor hogere wisselingen in de stroom ontstaan. Met behulp van de geautomatiseerde analysesoftware die bij het instrument wordt geleverd (Tabel van materialen), wordt I-Respons gemeten en geregistreerd in termen van biosensing units (BU)37. I-Respons wordt gedefinieerd als de verandering in de stroom (I) door de biosensorchip gemeten in real-time bij de interactie van het geïmmobiliseerde doelwit met de analyt. De geautomatiseerde analysesoftware van het VBO kan zowel de I-Respons als de C-Respons op dynamische interactiegebeurtenissen analyseren, waarbij de C-Respons de veranderingen in de capaciteit (C) registreert. De variaties in zowel de I-Respons als de C-Respons komen direct overeen met de fractie van gebonden analyt en kunnen verder worden geanalyseerd om KD-waarden te genereren. De standaardvoorkeur van de geautomatiseerde analysesoftware is I-Response.
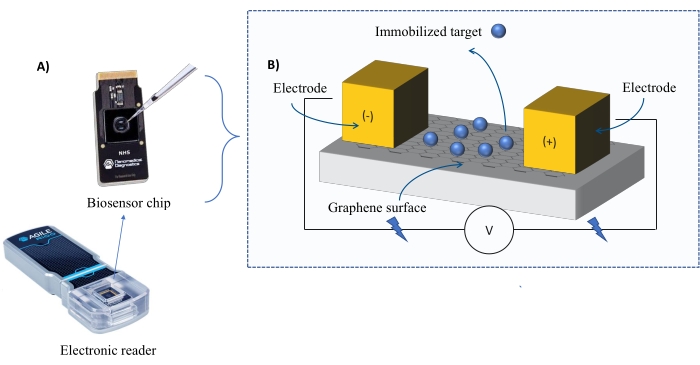
Figuur 1: Overzicht van de experimentele opstelling. (A) Op grafeen gebaseerde chip en een elektronische lezer. (B) Een overzicht van de chipcomponenten. De chip is bevestigd aan twee elektroden die stroom leveren aan het systeem. Het oppervlak van de chip is bedekt met grafeen, dat bij activering het doelwit kan binden. Klik hier om een grotere versie van deze figuur te bekijken.
Methodologie:
In eerste instantie wordt de geactiveerde biosensorchip in het FEB-apparaat ingebracht (figuur 1), gevolgd door de uitvoering van de onderstaande stappen: (1) Kalibratie: Het experiment begint met systeemkalibratie met behulp van 1x fosfaat-gebufferde zoutoplossing (PBS; pH = 7,4) om de basislijn equilibratierespons te creëren. (2) Associatie: De analyt wordt in de chip geïntroduceerd en de I-respons wordt gemonitord totdat de bindingsverzadiging is bereikt. (3) Dissociatie: De analyt wordt gedissocieerd met behulp van 1x PBS. (4) Regeneratie: Restanten van de analyt worden verwijderd met behulp van 1x PBS. (5) Wassen: In totaal worden vijf wasbeurten uitgevoerd met behulp van 1x PBS voor het grondig verwijderen van de gebonden en ongebonden analyten uit de chip.
Analyse:
Data-analyse wordt uitgevoerd met behulp van de volledig geautomatiseerde software die bij het instrument wordt geleverd. De geautomatiseerde analysesoftware genereert een Hill fit plot met eenKD-waarde . De Hill fit plot beschrijft de associatie van een analyt met het doeleiwit als functie van analytconcentraties. De concentratie waarbij een half-maximale respons wordt bereikt, is evenredig met de KD-waarde . Een lageKD-waarde vertegenwoordigt een hoge bindingsaffiniteit en vice versa.
Om de gegevens verkregen uit het VBO-experiment te valideren, worden I-responsen geëxtraheerd uit elk uitleespunt voor elke analytconcentratie met behulp van de data review/export software en kunnen ze worden geëxporteerd naar andere statistische analysesoftware (zie Tabel met materialen) zoals hieronder uitgelegd.
