Biomolekylære interaksjoner:
Proteiner er essensielle deler av organismer og deltar i mange molekylære veier som cellemetabolisme, cellestruktur, cellesignalering, immunresponser, celleadhesjon og mer. Mens noen proteiner utfører sin funksjon (er) uavhengig, samhandler de fleste proteiner med andre proteiner ved hjelp av et bindende grensesnitt for å koordinere riktig biologisk aktivitet1.
Biomolekylære interaksjoner kan hovedsakelig klassifiseres basert på de distinkte strukturelle og funksjonelle egenskapene til proteiner involvert2, for eksempel basert på proteinoverflatene, den komplekse stabiliteten eller utholdenheten til interaksjoner3. Identifisering av essensielle proteiner og deres roller i biomolekylære interaksjoner er avgjørende for å forstå biokjemiske mekanismer på molekylærnivå4. For tiden er det forskjellige tilnærminger for å oppdage disse interaksjonene5: in vitro6, i silico7, i levende celler8, ex vivo9 og in vivo10 med hver har sine egne styrker og svakheter.
In vivo-analysene utføres ved hjelp av hele dyret som et eksperimentelt verktøy11, oghan eks vivo-analyser utføres på vevsekstrakter eller hele organer (f.eks. hjerte, hjerne, lever) i et kontrollert eksternt miljø ved å gi minimale endringer i naturlige forhold. Den vanligste anvendelsen av in vivo– og ex vivo-studier er å evaluere farmakokinetikk, farmakodynamikk og toksisitetseffekter av potensielle farmakologiske midler før menneskelige studier ved å sikre deres generelle sikkerhet og effekt12.
Biomolekylære interaksjoner kan også påvises i levende celler. Imaging levende celler tillater oss å observere dynamiske interaksjoner som de utfører reaksjonene av en bestemt biokjemisk vei13. Videre kan deteksjonsteknikker, for eksempel bioluminescens eller fluorescensresonansenergioverføring, gi informasjon om hvor og når disse interaksjonene skjer i celle14. Selv om deteksjon i levende celler gir viktige detaljer, er disse deteksjonsmetodene avhengige av optikk og etiketter, noe som kanskje ikke gjenspeiler den opprinnelige biologien; de er også mindre kontrollerte enn in vitro-metoder og krever spesialisert kompetanse for å utføre15.
I silico beregningsmetoder brukes primært til storskala screening av målmolekyler før in vitro-forsøkene. Beregningsmessige prediksjonsmetoder, databaserte databaser, molekylær dokking, kvantitative strukturaktivitetsrelasjoner og andre molekylære dynamikksimuleringsmetoder er blant de veletablerte i silicoverktøy 16. Sammenlignet med arbeidskrevende eksperimentelle teknikker, kan in silico-verktøyene enkelt gjøre spådommer med høy følsomhet, men med redusert nøyaktighet i prediktiv ytelse17.
In vitro-analyser utføres med mikroorganismer eller biologiske molekyler utenfor deres standard biologiske kontekst. Skildring av biomolekylære interaksjoner gjennom in vitro-metoder er avgjørende for å forstå proteinfunksjoner og biologien bak det komplekse nettverket av cellefunksjon. Den foretrukne analysemetodikken velges i henhold til proteinets iboende egenskaper, kinetiske verdier og modusen og intensiteten av interaksjoner 18,19.
Hsp90/Cdc37-interaksjonen:
Chaperone-kinase-banen, som forbinder Hsp90 og Cdc37, er et lovende terapeutisk mål i tumorbiologi20. Hsp90 spiller en sentral rolle i cellesykluskontroll, proteinmontering, celleoverlevelse og signalveier. Proteiner som er avhengige av Hsp90 for deres funksjoner, leveres til Hsp90 for kompleksitet gjennom en medprest, for eksempel Cdc37. Hsp90/Cdc37-komplekset styrer foldingen av de fleste proteinkinaser og fungerer som et knutepunkt for en rekke intracellulære signalnettverk21. Det er et lovende anti-tumor mål på grunn av sin forhøyede uttrykk i ulike maligniteter, inkludert akutt myeloblastisk leukemi, multippel myeloma, og hepatocellulært karsinom22,23.
Vanlige in vitro biomolekylære interaksjonsdeteksjonsteknikker
Co-immunoprecipitation (co-IP) er en teknikk som er avhengig av antigen-antistoff spesifisitet for å identifisere biologisk relevante interaksjoner24. Den største ulempen ved denne metoden er dens manglende evne til å oppdage interaksjoner med lav affinitet og kinetiske verdier24. Biofysiske metoder som isotermisk titreringskalorimetri (ITC), overflateplasmonresonans (SPR), biolayer interferometri (BLI) og FEB-teknologi foretrekkes for å bestemme kinetiske verdier.
ITC er en biofysisk deteksjonsmetode basert på bestemmelse av bindende energi sammen med en komplett termodynamikkanalyse for å karakterisere biomolekylære interaksjoner25. Den viktigste fordelen med ITC er at det ikke krever merking eller fiksering av målproteinet. De viktigste vanskelighetene som ITC møter er den høye konsentrasjonen av målprotein som kreves for ett eksperiment og vanskeligheten med å analysere ikke-kovalente komplekser på grunn av små bindende entalpier26. Både SPR og BLI er etikettfrie biofysiske teknikker som er avhengige av immobilisering av målmolekylet på sensoroverflaten, etterfulgt av påfølgende injeksjoner av analytten over det immobiliserte målet27,28. I SPR måles endringer i brytningsindeksen under biomolekylære interaksjoner27; I BLI registreres interferensen i reflektert lys i sanntid som en endring i bølgelengden som en funksjon av tid28. Både SPR og BLI deler felles fordeler ved å tilby høy spesifisitet, følsomhet og deteksjonsevne29. I begge metoder er målproteinet immobilisert på biosensoroverflater, og derfor kan det være noe tap av målets opprinnelige konformasjon, noe som gjør det vanskelig å diskriminere mellom spesifikke kontra ikke-spesifikke interaksjoner30. BLI bruker dyre engangs fiberoptiske biosensorer for å immobilisere målet, og er derfor en kostbar teknikk31. Sammenlignet med disse veletablerte biomolekylære deteksjonsverktøyene, tilbyr FEB-teknologi en pålitelig og etikettfri plattform ved å bruke lave nanomolarkonsentrasjoner for biomolekylær deteksjon i sanntid med kinetisk karakterisering. FEB-teknologien overvinner også de boblende utfordringene i ITC og er mer kostnadseffektiv sammenlignet med SPR eller BLI.
Den felteffekttransistor (FET) basert biosensorer er et fremvoksende felt for å oppdage biomolekylære interaksjoner ved å tilby varierte biomedisinske applikasjoner. I FET-systemet blir mål immobilisert til biosensorbrikkene og interaksjoner oppdages ved endringer i ledning32. Den unike egenskapen som skal vurderes i utviklingen av en effektiv elektronisk biosensor er de fysisk-kjemiske egenskapene som den halvledende naturen og kjemisk stabilitet av beleggmaterialet som brukes til å fremstille sensoroverflaten33. Konvensjonelle materialer som silisium som brukes til FET har begrenset følsomheten til sensorer fordi det krever oksidlag smurt mellom transistorkanalen og et spesifikt miljø for riktig funksjon34. Videre er silisiumtransistorer følsomme for høye saltmiljøer, noe som gjør det vanskelig å måle biologiske interaksjoner i sitt naturlige miljø. Den grafenbaserte biosensoren presenteres som et alternativ, da den tilbyr utmerket kjemisk stabilitet og elektrisk felt. Siden grafen er et enkelt atomlag av karbon, er det både ekstremt følsomt som en halvleder og kjemisk kompatibel med biologiske løsninger; begge disse egenskapene er ønskelige å generere kompatible elektroniske biosensorer35. Det bemerkelsesverdige ultrahøye lastepotensialet til biomolekyler som tilbys av grafenbelagte biosensorer fører til utvikling av grafenbaserte biosensorer FEB-teknologi.
Prinsipp for FEB-teknologi: FEB er en etikettfri biomolekylær deteksjonsteknikk som måler den elektriske strømmen gjennom grafenbiosensoren som bindingsmålene immobiliseres til. Interaksjoner mellom det immobiliserte proteinet og analytten resulterer i endringer i strøm som overvåkes i sanntid, noe som muliggjør nøyaktige kinetiske målinger36.
Instrumentering: FEB-systemet består av en grafenfelteffekttransistorsensor (gFET) og en elektronisk leser som bruker konstant spenning gjennom hele eksperimentet (figur 1). Analytten påføres i løsning på målproteinet immobilisert på biosensoroverflaten. Når en interaksjon oppstår, måles og registreres en endring i strømmen i sanntid. Etter hvert som analyttkonsentrasjonen øker, vil brøkdelen av bundet analytt også øke, noe som forårsaker høyere vekslinger i strømmen. Ved hjelp av den automatiserte analyseprogramvaren som følger med instrumentet (Materialfortegnelse), måles og registreres I-Response når det gjelder biosensing-enheter (BU)37. I-Response er definert som endringen i den nåværende (I) gjennom biosensorbrikken målt i sanntid etter samspillet mellom det immobiliserte målet med analytten. Feb automatisert analyse programvare kan analysere både I-Response og C-Response til dynamiske interaksjon hendelser, der C-Response registrerer endringene i kapasitans (C). Variasjonene i både I-Response og C-Response samsvarer direkte med brøkdelen av bundet analytt og kan analyseres ytterligere for å generere KD-verdier . Standardinnstillingen for den automatiserte analyseprogramvaren er I-Response.
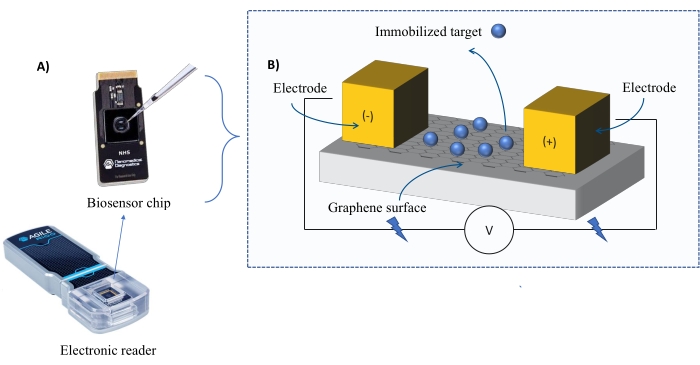
Figur 1: Oversikt over det eksperimentelle oppsettet. (A) Grafenbasert brikke og en elektronisk leser. (B) En oversikt over brikkekomponentene. Brikken er festet til to elektroder som leverer strøm til systemet. Overflaten på brikken er dekket med grafen, som når den aktiveres kan binde målet. Klikk her for å se en større versjon av denne figuren.
Metodikk:
I utgangspunktet settes den aktiverte biosensorbrikken inn i FEB-enheten (figur 1) etterfulgt av utførelsen av trinnene som er beskrevet nedenfor: (1) Kalibrering: Eksperimentet starter med systemkalibrering ved hjelp av 1x fosfatbufret saltvann (PBS; pH = 7,4) for å skape baseline likevektsrespons. (2) Forening: Analytten innføres i brikken, og I-Response overvåkes til bindingsmetningen er nådd. (3) Dissosiasjon: Analytten er dissosiert ved hjelp av 1x PBS. (4) Regenerering: Rester av analytten fjernes ved hjelp av 1x PBS. (5) Vasking: Totalt fem vasker utføres ved hjelp av 1x PBS for grundig fjerning av de bundne og ubundne analyttene fra brikken.
Analyse:
Dataanalyse utføres ved hjelp av den helautomatiske programvaren som følger med instrumentet. Den automatiserte analyseprogramvaren genererer en Hill fit-tomt med en KD-verdi . Hill fit plott beskriver foreningen av en analytt til målproteinet som en funksjon av analyttkonsentrasjoner. Konsentrasjonen der en halvmaksimal respons oppnås er proporsjonal med KD-verdien . En lav KD-verdi representerer høy bindingsaffinitet og omvendt.
For å validere dataene som er hentet fra FEB-eksperimentet, blir I-Responses hentet fra hvert avlesningspunkt for hver analyttkonsentrasjon ved hjelp av datagjennomgangs-/eksportprogramvaren og kan eksporteres til annen statistisk analyseprogramvare (se Tabell over materialer) som forklart nedenfor.
