يزداد تسلل الدهون في العضلات الهيكلية ، والمعروف باسم myosteatosis ، مع السمنة والشيخوخة. يرتبط Myosteatosis سلبا بكتلة العضلات وقوتها ومع حساسية الأنسولين1. علاوة على ذلك ، تشير الدراسات الحديثة إلى أنه يمكن استخدام درجة تنخر العظم العضلي كعامل تنبؤي لحالات أخرى مثل أمراض القلب والأوعية الدموية2 أو مرض الكبد الدهني غير الكحولي3 أو السرطان4. يمكن أن تتراكم الدهون في العضلات الهيكلية بين ألياف العضلات كدهون خارج الخلية أو داخل الألياف ، مثل الدهون داخل الخلايا (IMCLs). يتم تخزين IMCLs في الغالب كدهون ثلاثية في قطرات الدهون (LDs) التي تستخدم كوقود استقلابي أثناء التمرين البدني 5,6. ومع ذلك ، عندما يتجاوز عرض الدهون الطلب ، أو عندما تصبح الميتوكوندريا مختلة وظيفيا ، فإن IMCLs سوف تكون متورطة في مقاومة الأنسولين العضلي ، كما هو موضح في الأفراد غير الأصحاء والسمنة الأيضية وفي مرضى السكري من النوع 27. ومن المثير للاهتمام ، أن رياضيي التحمل لديهم مستويات مماثلة ، إن لم تكن أعلى ، من IMCLs لتلك الموجودة في المرضى الذين يعانون من السمنة المفرطة مع داء السكري من النوع 2 ، مع الحفاظ على حساسية عالية للأنسولين. توصف هذه الظاهرة بأنها “مفارقة الرياضي”8,9 ، ويتم تفسيرها من خلال تقييم أكثر دقة ل LDs العضلات ، المتعلقة بحجمها وكثافتها وتوطينها وديناميكياتها وتكوين أنواع الدهون.
أولا ، يرتبط حجم LD عكسيا بحساسية الأنسولين واللياقة البدنية10,11. في الواقع ، تظهر LDs الأصغر مساحة سطح أكبر نسبيا لعمل الليباز ، وبالتالي ، من المحتمل أن يكون لها قدرة أكبر على تعبئة الدهون12. ثانيا ، تلعب كثافة LD (العدد / السطح) دورا مثيرا للجدل في عمل الأنسولين 8,10 ؛ ومع ذلك ، يبدو أنه يزداد في الرياضيين. ثالثا ، يعد التوطين تحت الخلوي ل LDs مهما ، لأن LDs الموجودة أسفل الغشاء السطحي مباشرة (تحت الساركوليمال أو المحيطي) تمارس تأثيرا أكثر ضررا على حساسية الأنسولين من تلك المركزية 8,9,13. وتوفر هذه الأخيرة الوقود للميتوكوندريا المركزية، التي لديها نشاط تنفسي أكبر وأكثر تخصصا لتلبية الطلب المرتفع على الطاقة اللازم للانكماش14. وعلى النقيض من ذلك، توفر LDs الطرفية الميتوكوندريا تحت الساركوليمالية، التي تشارك في العمليات المتعلقة بالأغشية8. أخيرا ، بالإضافة إلى الدهون الثلاثية ، قد تكون الدهون المعقدة المحددة داخل العضلات أكثر ضررا من غيرها. على سبيل المثال ، قد يتراكم diacylglycerol و acyl-CoA طويل السلسلة والسيراميد في العضلات عندما يكون معدل دوران الدهون الثلاثية منخفضا ، مما يضعف إشارات الأنسولين 9,15. بالعودة إلى “مفارقة الرياضي” ، فإن رياضيي التحمل لديهم عدد كبير من LDs المركزية الأصغر حجما مع معدلات دوران مرتفعة في ألياف النوع الأول (المؤكسد) ، في حين أن مرضى السمنة والسكري لديهم LDs محيطية أكبر مع معدلات دوران منخفضة في ألياف النوع الثاني (السكري)8،15،16. بالإضافة إلى دورها في تخزين الطاقة وإطلاقها ، يمكن أن تعمل LDs عبر الأحماض الدهنية المشتقة (FA) وبروتين المعطف (perilipin 5) أيضا كلاعبين حاسمين يشاركون في التنظيم النسخي لأكسدة FA والتكوين الحيوي للميتوكوندريا8. بسبب آثارها الحاسمة في علم وظائف الأعضاء والفيزيولوجيا المرضية ، هناك ما يبرر إجراء دراسات متعمقة حول ديناميكيات ووظائف LDs.
على الرغم من وجود العديد من التقنيات لدراسة IMCLs ، إلا أنها ليست جميعها مناسبة لتحديد حجم LD وكثافته وتوزيعه بدقة بطريقة خاصة بالألياف. على سبيل المثال ، يوفر تقييم IMCLs بواسطة التحليل الطيفي بالرنين المغناطيسي ، على الرغم من كونه غير غازي ، مستوى من الدقة لا يكفي لدراسة حجم LDs وموقعها الدقيق داخل الألياف ، وليس من نوع الألياف المحدد17,18. وبالمثل ، لا يمكن للتقنيات الكيميائية الحيوية التي يتم إجراؤها على متجانسات العضلات بأكملها19 تقييم موقع وحجم الدهون. وبالتالي ، فإن الطريقة الأكثر ملاءمة لتحليل مورفولوجيا LD وموقعها هي المجهر الإلكتروني للإرسال الكمي13 ، ولكن هذه التقنية مكلفة وتستغرق وقتا طويلا. لذلك ، برز التصوير الفلوري البؤري على المستحضرات ذات الأصباغ مثل Oil Red O (ORO) 20,21 أو monodansylpentane (MDH)22 أو Bodipy 23,24,25 كأفضل أداة لهذه الدراسات.
هنا ، يتم وصف بروتوكول كامل ، بما في ذلك أخذ عينات الأنسجة ومعالجتها ، وتلطيخ Bodipy ، والحصول على الصور البؤرية وتحليلها لتحديد حجم LD وعددها وتوطينها في عمليات تبريد عضلات الفأر. نظرا لأن IMCLs لا يتم توزيعها بالتساوي بين الألياف المؤكسدة وتحلل السكر ، وينظم كل نوع من أنواع الألياف ديناميكيات LD بشكل مختلف ، يجب أن تكون دراسة IMCLs محددة من نوع الألياف 16,25,26,27. لذلك ، يستخدم هذا البروتوكول التألق المناعي على الأقسام التسلسلية لتحديد متساوي (أشكال) سلسلة الميوسين الثقيلة (MyHC) التي تعبر عنها كل ألياف. ميزة أخرى لهذا البروتوكول هي المعالجة المتزامنة ل glycolytic (الباسطة digitorum longus ، EDL) وعضلة مؤكسدة (وحيد) موضوعة جنبا إلى جنب قبل التجميد (الشكل 1). هذه المعالجة المتزامنة لا توفر الوقت فحسب ، بل تتجنب أيضا التباين بسبب المعالجة المنفصلة للعينات.
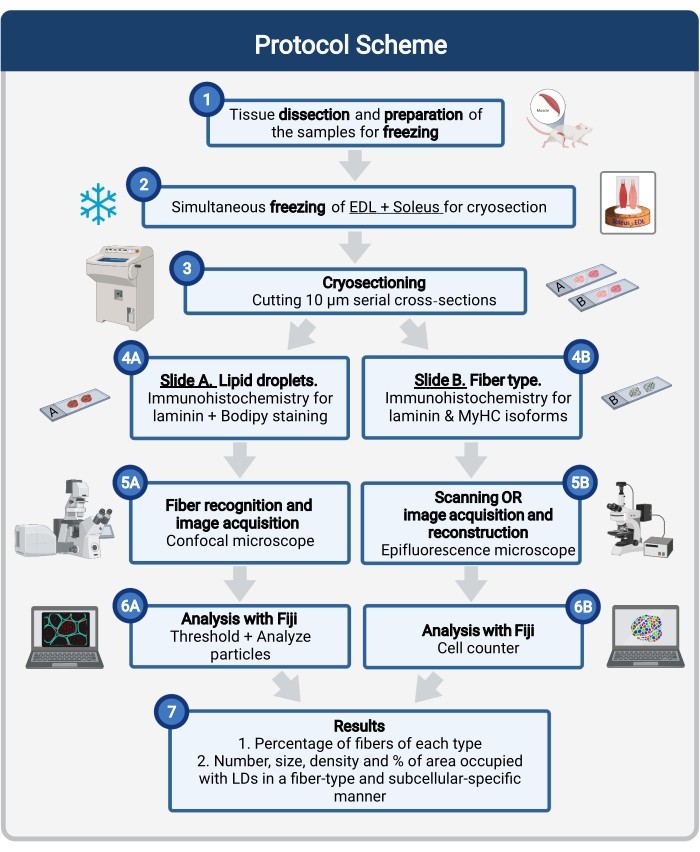
الشكل 1: نظرة عامة تخطيطية على الإجراء. بعد تشريح العضلات (1) ، يتم تحضير العضلات المختارة ذات الحجم المماثل وتجميدها معا (2). يتم الحصول على مقاطع عرضية تسلسلية من 10 ميكرومتر باستخدام cryostat ومثبتة مباشرة على شرائح الالتصاق (3). من شريحتين تسلسليتين ، الأولى (4A) مناعية للأمينين وملطخة ب Bodipy للتعرف على LDs والثانية (4B) ملطخة بالمناعة بالأجسام المضادة ضد MyHCs للتعرف على أنواع الألياف العضلية. يتم الحصول على الصور باستخدام مجهر متحد البؤرة ل Bodipy (5A) ومجهر epifluorescence لأنواع ألياف العضلات (5B). يتم تحليل الصور في فيجي من خلال تطبيق عتبة وتحديد كمية الجسيمات (6A) للحصول على العدد ومتوسط الحجم والكثافة والنسبة المئوية من المساحة الإجمالية التي تشغلها LDs (7) أو خلايا العد (6B) للحصول على النسبة المئوية للألياف من كل نوع في القسم (7). الاختصارات: LDs = قطرات الدهون. EDL = الباسطة الرقمية طويلة ؛ MyHCs = الميوسين سلسلة متساوية الثقيلة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
