Skeletspier lipide-infiltratie, bekend als myosteatose, neemt toe met obesitas en veroudering. Myosteatose is negatief gecorreleerd met spiermassa en kracht en met insulinegevoeligheid1. Bovendien geven recente studies aan dat de mate van myosteatose kan worden gebruikt als een prognostische factor voor andere aandoeningen zoals hart- en vaatziekten2, niet-alcoholische leververvetting3 of kanker4. Lipiden kunnen zich ophopen in skeletspieren tussen spiervezels als extramyocellulaire lipiden of in de vezels, als intramyocellulaire lipiden (IMCLs). IMCLs worden voornamelijk opgeslagen als triglyceriden in lipidedruppels (LD’s) die worden gebruikt als metabole brandstof tijdens lichaamsbeweging 5,6. Wanneer het lipidenaanbod echter de vraag overtreft, of wanneer mitochondriën disfunctioneel worden, zullen IMCLs betrokken zijn bij spierinsulineresistentie, zoals gezien bij metabolisch ongezonde, zwaarlijvige personen en bij type 2 diabetespatiënten7. Intrigerend genoeg hebben duursporters vergelijkbare, zo niet hogere, niveaus van IMCLs als die gevonden bij zwaarlijvige patiënten met type 2 diabetes mellitus, met behoud van een hoge insulinegevoeligheid. Dit fenomeen wordt beschreven als de “atleetparadox”8,9 en wordt verklaard door een meer genuanceerde beoordeling van spier-LD’s, gerelateerd aan hun grootte, dichtheid, lokalisatie, dynamiek en samenstelling van lipidesoorten.
Ten eerste is de LD-grootte omgekeerd gecorreleerd aan insulinegevoeligheid en fysieke fitheid10,11. In feite vertonen kleinere LD’s een relatief groter oppervlak voor lipasewerking en hebben ze dus mogelijk een groter vermogen om lipiden te mobiliseren12. Ten tweede speelt LD-dichtheid (getal/oppervlak) een controversiële rol bij insulinewerking 8,10; toch lijkt het bij sporters toe te nemen. Ten derde is de subcellulaire lokalisatie van LD’s belangrijk, omdat LD’s net onder het oppervlaktemembraan (subsarcolemmaal of perifeer) een schadelijker effect uitoefenen op de insulinegevoeligheid dan centrale 8,9,13. Deze laatste leveren brandstof aan centrale mitochondriën, die een grotere ademhalingsactiviteit hebben en meer gespecialiseerd zijn om te voldoen aan de hoge energievraag die nodig is voor contractie14. Perifere LD’s leveren daarentegen subsarcolemmale mitochondriën, die betrokken zijn bij membraangerelateerde processen8. Ten slotte kunnen, naast triglyceriden, specifieke complexe lipiden in de spier schadelijker zijn dan andere. Diacylglycerol, lange-keten acyl-CoA en ceramiden kunnen zich bijvoorbeeld ophopen in spieren wanneer de triglyceridenomloopsnelheid laag is, waardoor insuline wordt aangetast die 9,15 signaleert. Terugkerend naar de “paradox van de atleet”, hebben duursporters een groot aantal kleinere centrale LD’s met verhoogde omloopsnelheden in type I (oxidatieve) vezels, terwijl zwaarlijvige en diabetische patiënten grotere perifere LD’s hebben met lage omloopsnelheden in type II (glycolytische) vezels 8,15,16. Naast hun rol in energieopslag en -afgifte, kunnen LDs via afgeleide vetzuren (FA) en een vachteiwit (perilipin 5) ook functioneren als kritische spelers die betrokken zijn bij de transcriptionele regulatie van FA-oxidatie en mitochondriale biogenese8. Vanwege hun cruciale implicaties in de fysiologie en pathofysiologie, zijn diepgaande studies over de dynamiek en functies van LDs gerechtvaardigd.
Hoewel er verschillende technieken zijn om IMCLs te bestuderen, zijn ze niet allemaal geschikt om de LD-grootte, -dichtheid en -distributie op een vezelspecifieke manier nauwkeurig te kwantificeren. Bijvoorbeeld, de beoordeling van IMCLs door magnetische resonantiespectroscopie, hoewel niet-invasief, biedt een resolutieniveau dat niet voldoende is om de grootte en precieze locatie van LDs in de vezel te bestuderen, en het is niet vezeltypespecifiek17,18. Evenzo kunnen biochemische technieken uitgevoerd op homogene homogenen van de hele spieren19 de locatie en grootte van lipiden niet beoordelen. Bijgevolg is de meest geschikte methode om LD-morfologie en locatie te analyseren kwantitatieve transmissie-elektronische microscopie13, maar deze techniek is duur en tijdrovend. Daarom is confocale fluorescentiebeeldvorming op preparaten met kleurstoffen zoals Oil Red O (ORO)20,21, monodansylpentane (MDH)22 of Bodipy 23,24,25 naar voren gekomen als het beste hulpmiddel voor deze studies.
Hier wordt een compleet protocol beschreven, inclusief weefselbemonstering en -verwerking, Bodipy-kleuring en confocale beeldacquisitie en -analyse om de LD-grootte, het aantal en de lokalisatie in cryosecties van muizenspieren te kwantificeren. Aangezien IMCLs niet gelijkmatig verdeeld zijn over oxidatieve en glycolytische vezels, en elk vezeltype de LD-dynamiek anders reguleert, moet de studie van IMCLs vezeltypespecifiekzijn 16,25,26,27. Daarom gebruikt dit protocol immunofluorescentie op seriële secties om myosine zware keten (MyHC) isovorm (en) uitgedrukt door elke vezel te identificeren. Een ander voordeel van dit protocol is de gelijktijdige verwerking van een glycolytische (extensor digitorum longus, EDL) en een oxidatieve (soleus) spier die naast elkaar worden geplaatst voor het bevriezen (figuur 1). Deze gelijktijdige verwerking bespaart niet alleen tijd, maar voorkomt ook variabiliteit door afzonderlijke verwerking van de monsters.
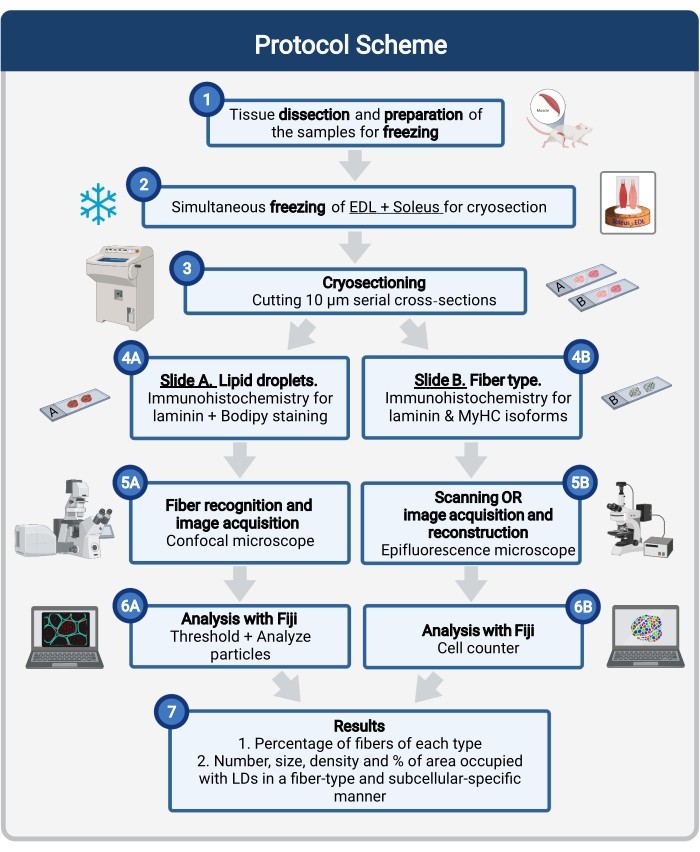
Figuur 1: Schematisch overzicht van de procedure. Na spierdissectie (1) worden geselecteerde spieren van vergelijkbare grootte voorbereid en samen ingevroren (2). Seriële dwarsdoorsneden van 10 μm worden verkregen met behulp van een cryostaat en direct gemonteerd op adhesieglaasjes (3). Van twee seriële dia’s is de eerste (4A) immunoactief gelabeld voor laminine en gekleurd met Bodipy om LDs te herkennen en de tweede (4B) is immunostained met antilichamen tegen MyHC’s voor de herkenning van spiervezeltypen. Beelden worden verkregen met behulp van een confocale microscoop voor Bodipy (5A) en een epifluorescentiemicroscoop voor spiervezeltypen (5B). Afbeeldingen worden in Fiji geanalyseerd door een drempel toe te passen en deeltjes (6A) te kwantificeren om het aantal, de gemiddelde grootte, de dichtheid en het percentage van het totale gebied bezet door LDs (7) of telcellen (6B) te verkrijgen om het percentage vezels van elk type in de sectie (7) te verkrijgen. Afkortingen: LDs = lipidedruppels; EDL = extensor digitorum longus; MyHC’s = myosine zware keten isovormen. Klik hier om een grotere versie van deze figuur te bekijken.
