L’infiltrazione lipidica del muscolo scheletrico, nota come miosteatosi, aumenta con l’obesità e l’invecchiamento. La miosteatosi è correlata negativamente con la massa e la forza muscolare e con la sensibilità all’insulina1. Inoltre, studi recenti indicano che il grado di miosteatosi potrebbe essere utilizzato come fattore prognostico per altre condizioni come la malattia cardiovascolare2, la steatosi epatica non alcolica3 o il cancro4. I lipidi possono accumularsi nel muscolo scheletrico tra le fibre muscolari come lipidi extramiocellulari o all’interno delle fibre, come lipidi intramiocellulari (IMCL). Gli IMCL sono prevalentemente immagazzinati come trigliceridi in goccioline lipidiche (LD) che vengono utilizzate come combustibile metabolico durante l’esercizio fisico 5,6. Tuttavia, quando l’offerta lipidica supera la domanda, o quando i mitocondri diventano disfunzionali, gli IMCL saranno implicati nella resistenza all’insulina muscolare, come si vede in individui metabolicamente malsani e obesi e nei pazienti con diabete di tipo 27. Curiosamente, gli atleti di resistenza hanno livelli simili, se non più alti, di IMCL a quelli trovati nei pazienti obesi con diabete mellito di tipo 2, pur mantenendo un’elevata sensibilità all’insulina. Questo fenomeno è descritto come il “paradosso dell’atleta”8,9, ed è spiegato da una valutazione più sfumata degli LCD muscolari, relativi alle loro dimensioni, densità, localizzazione, dinamica e composizione delle specie lipidiche.
In primo luogo, la dimensione della LD è inversamente correlata alla sensibilità all’insulina e alla forma fisica10,11. In effetti, le LD più piccole mostrano una superficie relativamente maggiore per l’azione della lipasi e, quindi, potenzialmente hanno una maggiore capacità di mobilitare i lipidi12. In secondo luogo, la densità LD (numero/superficie) svolge un ruolo controverso nell’azione dell’insulina 8,10; tuttavia, sembra essere aumentato negli atleti. In terzo luogo, la localizzazione subcellulare delle LD è importante, poiché le LD situate appena sotto la membrana superficiale (subsarcolemmale o periferica) esercitano un effetto più deleterio sulla sensibilità all’insulina rispetto a quelle centrali 8,9,13. Questi ultimi forniscono carburante ai mitocondri centrali, che hanno una maggiore attività respiratoria e sono più specializzati per soddisfare l’elevata domanda di energia richiesta per la contrazione14. Al contrario, le LD periferiche forniscono mitocondri subsarcolemmali, che sono coinvolti nei processi correlati alla membrana8. Infine, oltre ai trigliceridi, specifici lipidi complessi all’interno del muscolo possono essere più deleteri di altri. Ad esempio, il diacilglicerolo, l’acil-CoA a catena lunga e le ceramidi possono accumularsi nel muscolo quando il tasso di turnover dei trigliceridi è basso, compromettendo così la segnalazione dell’insulina 9,15. Tornando al “paradosso dell’atleta”, gli atleti di resistenza hanno un numero elevato di LD centrali più piccoli con elevati tassi di turnover nelle fibre di tipo I (ossidative), mentre i pazienti obesi e diabetici hanno LD periferici più grandi con bassi tassi di turnover nelle fibre di tipo II (glicolitiche) 8,15,16. Oltre al loro ruolo nell’immagazzinamento e nel rilascio di energia, gli LCD tramite acidi grassi derivati (FA) e una proteina del mantello (perilipina 5) potrebbero anche funzionare come attori critici coinvolti nella regolazione trascrizionale dell’ossidazione fa e della biogenesi mitocondriale8. A causa delle loro implicazioni cruciali in fisiologia e fisiopatologia, sono giustificati studi approfonditi sulla dinamica e le funzioni dei LD.
Sebbene esistano diverse tecniche per studiare le IMCL, non sono tutte adatte a quantificare con precisione le dimensioni, la densità e la distribuzione dei LD in modo specifico per le fibre. Ad esempio, la valutazione delle IMCL mediante spettroscopia di risonanza magnetica, pur essendo non invasiva, offre un livello di risoluzione che non è sufficiente per studiare le dimensioni e la posizione precisa degli LCD all’interno della fibra, e non è specifico per il tipo di fibra17,18. Allo stesso modo, le tecniche biochimiche eseguite su omogeneizzati muscolari interi19 non possono valutare la posizione e le dimensioni dei lipidi. Di conseguenza, il metodo più adeguato per analizzare la morfologia e la posizione della LD è la microscopia elettronica a trasmissione quantitativa13, ma questa tecnica è costosa e richiede tempo. Pertanto, l’imaging a fluorescenza confocale su preparati con coloranti come Oil Red O (ORO)20,21, monodansylpentane (MDH)22 o Bodipy 23,24,25, è emerso come lo strumento migliore per questi studi.
Qui viene descritto un protocollo completo, che include il campionamento e l’elaborazione dei tessuti, la colorazione di Bodipy e l’acquisizione e l’analisi di immagini confocali per quantificare le dimensioni, il numero e la localizzazione di LD nelle criosezioni muscolari del topo. Poiché gli IMCL non sono distribuiti uniformemente tra fibre ossidative e glicolitiche e ogni tipo di fibra regola la dinamica LD in modo diverso, lo studio delle IMML deve essere specifico per il tipo di fibra 16,25,26,27. Pertanto, questo protocollo utilizza l’immunofluorescenza su sezioni seriali per identificare le isoforme a catena pesante della miosina (MyHC) espresse da ciascuna fibra. Un altro vantaggio di questo protocollo è l’elaborazione simultanea di un muscolo glicolitico (estensore digitorum longus, EDL) e di un muscolo ossidativo (soleo) posti fianco a fianco prima del congelamento (Figura 1). Questa elaborazione simultanea non solo consente di risparmiare tempo, ma evita anche la variabilità dovuta all’elaborazione separata dei campioni.
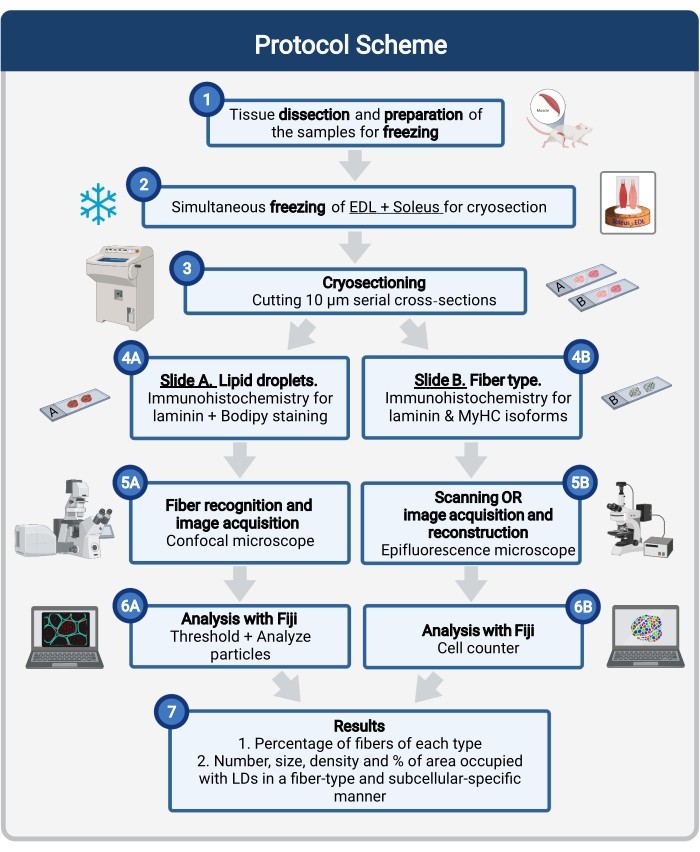
Figura 1: Panoramica schematica della procedura. Dopo la dissezione muscolare (1), i muscoli selezionati di dimensioni simili vengono preparati e congelati insieme (2). Le sezioni trasversali seriali di 10 μm sono ottenute utilizzando un criostato e montate direttamente su vetrini di adesione (3). Da due vetrini seriali, il primo (4A) è immunomarcato per la laminina e colorato con Bodipy per riconoscere gli LD e il secondo (4B) è immunocolorato con anticorpi contro i MyHC per il riconoscimento dei tipi di fibre muscolari. Le immagini vengono acquisite utilizzando un microscopio confocale per Bodipy (5A) e un microscopio a epifluorescenza per i tipi di fibre muscolari (5B). Le immagini vengono analizzate nelle Figi applicando una soglia e quantificando le particelle (6A) per ottenere il numero, la dimensione media, la densità e la percentuale dell’area totale occupata da LD (7) o celle di conteggio (6B) per ottenere la percentuale di fibre di ciascun tipo nella sezione (7). Abbreviazioni: LDs = goccioline lipidiche; EDL = estensore digitorum longus; MyHC = isoforme a catena pesante della miosina. Fare clic qui per visualizzare una versione più grande di questa figura.
