Skeletmuskel lipid infiltration, kendt som myosteatose, stiger med fedme og aldring. Myosteatose er negativt korreleret med muskelmasse og styrke og med insulinfølsomhed1. Desuden viser nylige undersøgelser, at graden af myosteatose kan anvendes som en prognostisk faktor for andre tilstande såsom hjerte-kar-sygdom2, ikke-alkoholisk fedtleversygdom3 eller kræft4. Lipider kan akkumuleres i skeletmuskulatur mellem muskelfibre som ekstramyocellulære lipider eller i fibrene, som intramyocellulære lipider (IMCLs). IMCL’er opbevares overvejende som triglycerider i lipiddråber (LD’er), der anvendes som metabolisk brændstof under fysisk træning 5,6. Men når lipidforsyningen overstiger efterspørgslen, eller når mitokondrier bliver dysfunktionelle, vil IMCLs være impliceret i muskelinsulinresistens, som det ses hos metabolisk usunde, overvægtige individer og hos type 2-diabetespatienter7. Spændende nok har udholdenhedsatleter lignende, hvis ikke højere, niveauer af IMCLs til dem, der findes hos overvægtige patienter med type 2-diabetes mellitus, samtidig med at de opretholder høj insulinfølsomhed. Dette fænomen beskrives som “atletens paradoks”8,9 og forklares ved en mere nuanceret vurdering af muskel-ID’er relateret til deres størrelse, tæthed, lokalisering, dynamik og lipidartssammensætning.
For det første er LD-størrelse omvendt korreleret med insulinfølsomhed og fysisk kondition10,11. Faktisk udviser mindre LD’er et relativt større overfladeareal til lipasevirkning og har således potentielt en større kapacitet til at mobilisere lipider12. For det andet spiller LD-densitet (tal/overflade) en kontroversiel rolle i insulinvirkning 8,10; alligevel ser det ud til at være øget hos atleter. For det tredje er den subcellulære lokalisering af LD’er vigtig, da LD’er placeret lige under overflademembranen (subsarcolemmal eller perifer) udøver en mere skadelig virkning på insulinfølsomheden end centrale 8,9,13. Sidstnævnte giver brændstof til centrale mitokondrier, som har en større respiratorisk aktivitet og er mere specialiserede til at imødekomme det høje energibehov, der kræves til sammentrækning14. I modsætning hertil leverer perifere LD’er subsarcolemmale mitokondrier, som er involveret i membranrelaterede processer8. Endelig, ud over triglycerider, kan specifikke komplekse lipider i musklen være mere skadelige end andre. For eksempel kan diacylglycerol, langkædet acyl-CoA og ceramider akkumulere i muskler, når triglyceridomsætningshastigheden er lav, hvilket forringer insulinsignalering 9,15. For at vende tilbage til “atletens paradoks” har udholdenhedsatleter et stort antal mindre centrale LD’er med forhøjede omsætningshastigheder i type I (oxidative) fibre, mens overvægtige og diabetespatienter har større perifere LD’er med lave omsætningshastigheder i type II (glykolytiske) fibre 8,15,16. Ud over deres rolle i energilagring og frigivelse kunne LD’er via afledte fedtsyrer (FA) og et pelsprotein (perilipin 5) også fungere som kritiske aktører involveret i transkriptionel regulering af FA-oxidation og mitokondriebiogenese8. På grund af deres afgørende implikationer i fysiologi og patofysiologi er dybtgående undersøgelser af LD’ers dynamik og funktioner berettiget.
Selvom der er flere teknikker til at studere IMCL’er, er de ikke alle egnede til nøjagtigt at kvantificere LD-størrelse, densitet og distribution på en fiberspecifik måde. For eksempel tilbyder vurderingen af IMCLs ved magnetisk resonansspektroskopi, selvom den ikke er invasiv, et opløsningsniveau, der ikke er tilstrækkeligt til at studere størrelsen og den præcise placering af LD’er i fiberen, og det er ikke fibertypespecifikt17,18. Ligeledes kan biokemiske teknikker udført på helmuskelhomogenater19 ikke vurdere placeringen og størrelsen af lipider. Følgelig er den mest passende metode til analyse af LD-morfologi og placering kvantitativ transmissionselektronisk mikroskopi13, men denne teknik er dyr og tidskrævende. Derfor har konfokal fluorescensbilleddannelse på præparater med farvestoffer som Oil Red O (ORO)20,21, monodansylpentan (MDH)22 eller Bodipy 23,24,25 vist sig som det bedste værktøj til disse undersøgelser.
Her beskrives en komplet protokol, herunder vævsprøvetagning og -behandling, Bodipy-farvning og konfokal billedoptagelse og analyse for at kvantificere LD-størrelse, antal og lokalisering i musemuskelkryosektioner. Da IMCL’er ikke er jævnt fordelt mellem oxidative og glykolytiske fibre, og hver fibertype regulerer LD-dynamik forskelligt, skal undersøgelsen af IMCLs være fibertypespecifik 16,25,26,27. Derfor bruger denne protokol immunfluorescens på serielle sektioner til at identificere myosin tung kæde (MyHC) isoform (er) udtrykt af hver fiber. En anden fordel ved denne protokol er samtidig behandling af en glykolytisk (extensor digitorum longus, EDL) og en oxidativ (soleus) muskel placeret side om side før frysning (figur 1). Denne samtidige behandling sparer ikke kun tid, men undgår også variabilitet på grund af separat behandling af prøverne.
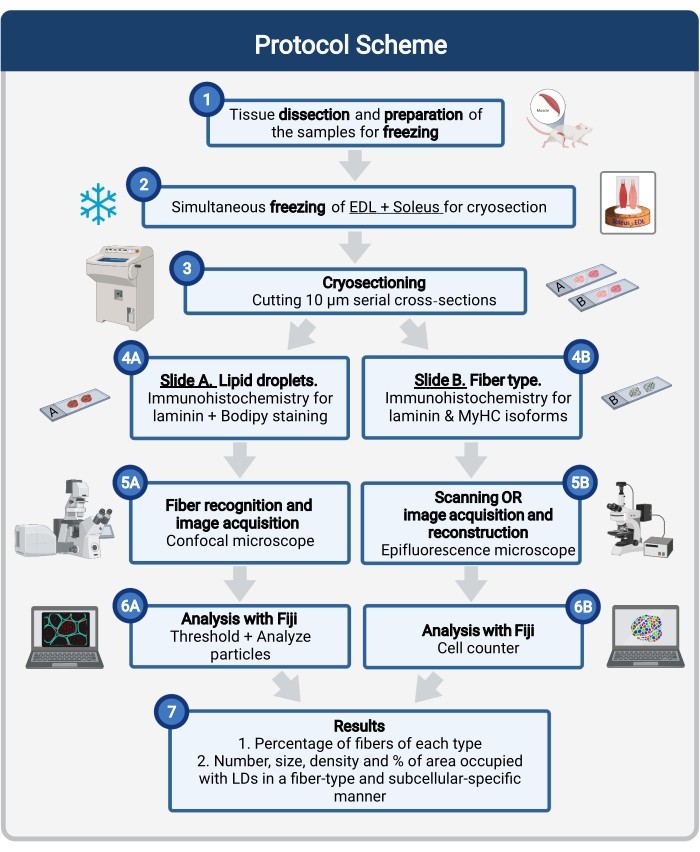
Figur 1: Skematisk oversigt over proceduren. Efter muskeldissektion (1) fremstilles udvalgte muskler af samme størrelse og fryses sammen (2). Serielle tværgående sektioner på 10 μm opnås ved anvendelse af en kryostat og monteres direkte på adhæsionsglas (3). Fra to serielle dias er den første (4A) immunmærket for laminin og farvet med Bodipy for at genkende LD’er, og den anden (4B) er immunostained med antistoffer mod MyHC’er til genkendelse af muskelfibertyper. Billeder erhverves ved hjælp af et konfokalt mikroskop til Bodipy (5A) og et epifluorescensmikroskop til muskelfibertyper (5B). Billeder analyseres i Fiji ved at anvende en tærskel og kvantificere partikler (6A) for at opnå antallet, gennemsnitsstørrelsen, densiteten og procentdelen af det samlede areal, der er optaget af LD’er (7) eller tælleceller (6B) for at opnå procentdelen af fibre af hver type i sektionen (7). Forkortelser: LD’er = lipiddråber; EDL = extensor digitorum longus; MyHC’er = myosin tunge kæde isoformer. Klik her for at se en større version af denne figur.
