Skjelettmuskel lipidinfiltrasjon, kjent som myosteatose, øker med fedme og aldring. Myosteatose er negativt korrelert med muskelmasse og styrke og med insulinfølsomhet1. Videre indikerer nyere studier at graden av myosteatose kan brukes som en prognostisk faktor for andre forhold som kardiovaskulær sykdom2, alkoholfri fettleversykdom3 eller kreft4. Lipider kan akkumuleres i skjelettmuskulatur mellom muskelfibre som ekstramyocellulære lipider eller i fibrene, som intramyocellulære lipider (IMCLer). IMCLer lagres hovedsakelig som triglyserider i lipiddråper (LDer) som brukes som metabolsk drivstoff under fysisk trening 5,6. Men når lipidforsyningen overstiger etterspørselen, eller når mitokondrier blir dysfunksjonelle, vil IMCLer bli implisert i muskelinsulinresistens, sett hos metabolsk usunne, overvektige individer og hos type 2 diabetespasienter7. Interessant nok har utholdenhetsutøvere lignende, om ikke høyere, nivåer av IMCLer til de som finnes hos overvektige pasienter med type 2 diabetes mellitus, samtidig som de opprettholder høy insulinfølsomhet. Dette fenomenet er beskrevet som “idrettsutøverens paradoks”8,9, og forklares av en mer nyansert vurdering av muskel-ID-er, relatert til deres størrelse, tetthet, lokalisering, dynamikk og lipidartsammensetning.
For det første er LD-størrelsen omvendt korrelert med insulinfølsomhet og fysisk form10,11. Faktisk viser mindre LP-er et relativt større overflateareal for lipasevirkning og har dermed potensielt større kapasitet til å mobilisere lipider12. For det andre spiller LD-tetthet (antall/overflate) en kontroversiell rolle i insulinvirkning 810; likevel ser det ut til å økes hos idrettsutøvere. For det tredje er subcellulær lokalisering av LP-er viktig, siden LP-er som ligger like under overflatemembranen (subsarcolemmal eller perifer) utøver en mer skadelig effekt på insulinfølsomhet enn sentrale 8,9,13. Sistnevnte gir drivstoff til sentrale mitokondrier, som har større åndedrettsaktivitet og er mer spesialiserte for å møte den høye energietterspørselen som kreves for sammentrekning14. Til sammenligning leverer perifere LDer subsarcolemmal mitokondrier, som er involvert i membranrelaterte prosesser8. Til slutt, utover triglyserider, kan spesifikke komplekse lipider i muskelen være mer skadelige enn andre. For eksempel kan diacylglyserol, langkjedet acyl-CoA og ceramider akkumuleres i muskel når triglyseridomsetningen er lav, og dermed svekke insulinsignalering 9,15. Tilbake til “idrettsutøverens paradoks”, utholdenhet idrettsutøvere har et høyt antall mindre sentrale ID-er med forhøyede omsetningshastigheter i type I (oksidative) fibre, mens overvektige og diabetikere har større perifere ID-er med lave omsetningshastigheter i type II (glykolytiske) fibre 8,15,16. I tillegg til deres rolle i energilagring og frigjøring, kan ID-er via avledede fettsyrer (FA) og et frakkprotein (perilipin 5) også fungere som kritiske aktører involvert i transkripsjonsreguleringen av FA-oksidasjon og mitokondriebiogenese8. På grunn av deres avgjørende implikasjoner i fysiologi og patofysiologi, er grundige studier av LDs dynamikk og funksjoner berettiget.
Selv om det finnes flere teknikker for å studere IMCLer, er de ikke alle egnet til å nøyaktig kvantifisere LD-størrelse, tetthet og distribusjon på en fiberspesifikk måte. For eksempel tilbyr vurderingen av IMCLer ved magnetisk resonansspektroskopi, mens den ikke er invasiv, et oppløsningsnivå som ikke er tilstrekkelig til å studere størrelsen og den nøyaktige plasseringen av LP-er i fiberen, og den er ikke fibertypespesifikk17,18. På samme måte kan biokjemiske teknikker utført på helmuskel homogenater19 ikke vurdere plasseringen og størrelsen på lipider. Følgelig er den mest tilstrekkelige metoden for å analysere LD morfologi og plassering kvantitativ overføring elektronisk mikroskopi13, men denne teknikken er dyr og tidkrevende. Derfor har konfekt fluorescensavbildning på preparater med fargestoffer som Oil Red O (ORO)20,21, monodansylpentane (MDH)22 eller Bodipy 23,24,25, dukket opp som det beste verktøyet for disse studiene.
Her beskrives en komplett protokoll, inkludert vevsprøvetaking og -behandling, Bodipy-farging og konfokal bildeanskaffelse og analyse for å kvantifisere LD-størrelse, antall og lokalisering i musemuskelroposer. Siden IMCLer ikke er jevnt fordelt mellom oksidative og glykolytiske fibre, og hver fibertype regulerer LD-dynamikken annerledes, må studien av IMCLer være fibertypespesifikk 16,25,26,27. Derfor bruker denne protokollen immunfluorescens på serielle seksjoner for å identifisere myosin heavy chain (MyHC) isoform(er) uttrykt av hver fiber. En annen fordel med denne protokollen er samtidig behandling av en glykolytisk (extensor digitorum longus, EDL) og en oksidativ (soleus) muskel plassert side om side før frysing (figur 1). Denne samtidige behandlingen sparer ikke bare tid, men unngår også variasjon på grunn av separat behandling av prøvene.
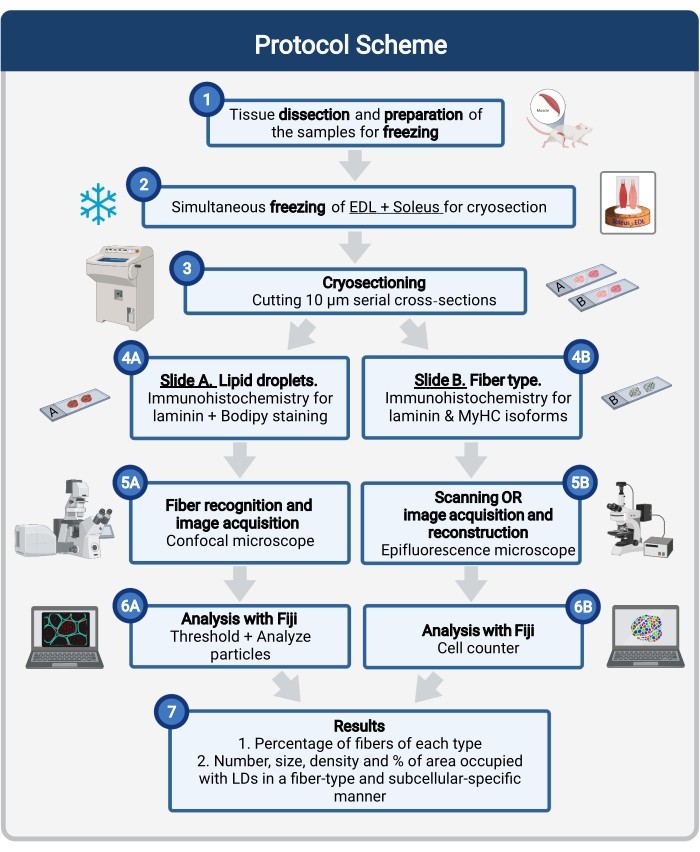
Figur 1: Skjematisk oversikt over prosedyren. Etter muskel disseksjon (1) fremstilles og fryses utvalgte muskler i tilsvarende størrelse sammen (2). Serielle tverrgående deler på 10 μm oppnås ved hjelp av en kryostat og monteres direkte på vedheftsskred (3). Fra to serielle lysbilder er den første (4A) immunmerket for laminin og farget med Bodipy for å gjenkjenne LD-er, og den andre (4B) er immunostert med antistoffer mot MyHCer for anerkjennelse av muskelfibertyper. Bilder er anskaffet ved hjelp av et konfokalt mikroskop for Bodipy (5A) og et epifluorescensmikroskop for muskelfibertyper (5B). Bilder analyseres i Fiji ved å bruke en terskel og kvantifiserende partikler (6A) for å oppnå tall, gjennomsnittlig størrelse, tetthet og prosentandel av det totale området okkupert av LP-er (7) eller telleceller (6B) for å oppnå prosentandelen fibre av hver type i seksjonen (7). Forkortelser: LDer = lipiddråper; EDL = extensor digitorum longus; MyHCs = myosin tunge kjede isoformer. Klikk her for å se en større versjon av denne figuren.
