Skelettmuskellipidinfiltration, känd som myosteatos, ökar med fetma och åldrande. Myosteatos är negativt korrelerad med muskelmassa och styrka och med insulinkänslighet1. Dessutom tyder nya studier på att graden av myosteatos kan användas som en prognostisk faktor för andra tillstånd som hjärt-kärlsjukdom2, alkoholfri fettleversjukdom3 eller cancer4. Lipider kan ackumuleras i skelettmuskulaturen mellan muskelfibrer som extramyocellulära lipider eller i fibrerna, som intramyocellulära lipider (IMCLs). IMCLs lagras främst som triglycerider i lipiddroppar (LDs) som används som metaboliskt bränsle under fysisk träning 5,6. Men när lipidutbudet överstiger efterfrågan, eller när mitokondrier blir dysfunktionella, kommer IMCLs att vara inblandade i muskelinsulinresistens, vilket ses hos metaboliskt ohälsosamma, överviktiga individer och hos typ 2-diabetespatienter7. Intressant nog har uthållighetsidrottare liknande, om inte högre, nivåer av IMCLs som de som finns hos överviktiga patienter med typ 2-diabetes mellitus, samtidigt som de bibehåller hög insulinkänslighet. Detta fenomen beskrivs som “idrottarens paradox”8,9 och förklaras av en mer nyanserad bedömning av muskel-LDs, relaterade till deras storlek, densitet, lokalisering, dynamik och lipidartsammansättning.
För det första är LD-storlek omvänt korrelerad till insulinkänslighet och fysisk kondition10,11. Faktum är att mindre LD uppvisar en relativt större ytarea för lipasverkan och därmed potentiellt har en större kapacitet att mobilisera lipider12. För det andra spelar LD-densitet (antal / yta) en kontroversiell roll i insulinverkan 8,10; ändå verkar det ökas hos idrottare. För det tredje är den subcellulära lokaliseringen av LDs viktig, eftersom LDs som ligger strax under ytmembranet (subsarcolemmal eller perifer) utövar en mer skadlig effekt på insulinkänsligheten än centrala 8,9,13. De senare ger bränsle till centrala mitokondrier, som har en större andningsaktivitet och är mer specialiserade för att möta det höga energibehovet som krävs för sammandragning14. Däremot levererar perifera LDs subsarcolemmal mitokondrier, som är involverade i membranrelaterade processer8. Slutligen, utöver triglycerider, kan specifika komplexa lipider i muskeln vara mer skadliga än andra. Till exempel kan diacylglycerol, långkedjig acyl-CoA och ceramider ackumuleras i muskler när triglyceridomsättningshastigheten är låg, vilket försämrar insulinsignalering 9,15. För att återgå till “idrottarens paradox” har uthållighetsidrottare ett stort antal mindre centrala LDs med förhöjda omsättningshastigheter i typ I (oxidativa) fibrer, medan överviktiga och diabetespatienter har större perifera LDs med låg omsättningshastighet i typ II (glykolytiska) fibrer 8,15,16. Förutom sin roll i energilagring och frisättning kan LDs via härledda fettsyror (FA) och ett pälsprotein (perilipin 5) också fungera som kritiska aktörer som är involverade i transkriptionell reglering av FA-oxidation och mitokondriell biogenes8. På grund av deras avgörande konsekvenser inom fysiologi och patofysiologi är fördjupade studier av LDs dynamik och funktioner motiverade.
Även om det finns flera tekniker för att studera IMCLs, är de inte alla lämpliga för att exakt kvantifiera LD-storlek, densitet och distribution på ett fiberspecifikt sätt. Till exempel erbjuder bedömningen av IMCLs genom magnetisk resonansspektroskopi, även om den är icke-invasiv, en upplösningsnivå som inte är tillräcklig för att studera storleken och den exakta placeringen av LD: er i fibern, och det är inte fibertypsspecifikt17,18. På samma sätt kan biokemiska tekniker som utförs på helmuskelhomogenater19 inte bedöma placeringen och storleken på lipider. Följaktligen är den mest adekvata metoden för att analysera LD-morfologi och plats kvantitativ överföring elektronisk mikroskopi13, men denna teknik är dyr och tidskrävande. Därför har konfokal fluorescensavbildning på preparat med färgämnen som Oil Red O (ORO)20,21, monodansylpentan (MDH)22 eller Bodipy 23,24,25 framstått som det bästa verktyget för dessa studier.
Här beskrivs ett komplett protokoll, inklusive vävnadsprovtagning och bearbetning, Bodipy-färgning och konfokal bildförvärv och analys för att kvantifiera LD-storlek, antal och lokalisering i musmuskelkryosektioner. Eftersom IMCLs inte är jämnt fördelade mellan oxidativa och glykolytiska fibrer, och varje fibertyp reglerar LD-dynamiken annorlunda, måste studien av IMCLs vara fibertypspecifik 16,25,26,27. Därför använder detta protokoll immunofluorescens på seriella sektioner för att identifiera myosin tunga kedjor (MyHC) isoform (er) uttryckt av varje fiber. En annan fördel med detta protokoll är samtidig behandling av en glykolytisk (extensor digitorum longus, EDL) och en oxidativ (soleus) muskel placerad sida vid sida före frysning (Figur 1). Denna samtidiga bearbetning sparar inte bara tid utan undviker också variabilitet på grund av separat bearbetning av proverna.
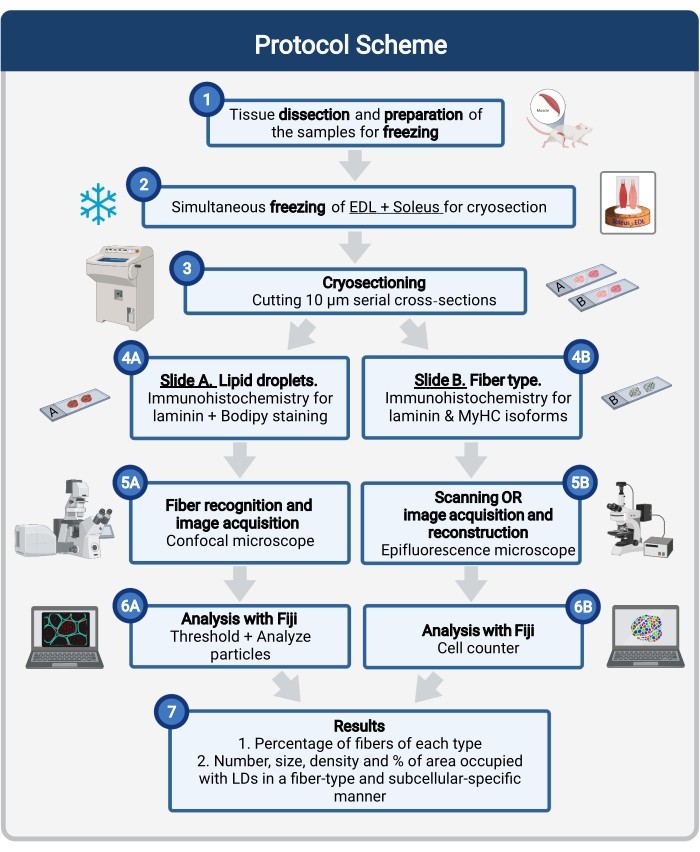
Figur 1: Schematisk översikt över proceduren. Efter muskeldissektion (1) förbereds och fryses utvalda muskler av liknande storlek (2). Seriella tvärsektioner på 10 μm erhålls med användning av en kryostat och monteras direkt på vidhäftningsglas (3). Från två seriella bilder är den första (4A) immunolabelerad för laminin och färgad med Bodipy för att känna igen LDs och den andra (4B) immunostained med antikroppar mot MyHC för erkännande av muskelfibertyper. Bilder förvärvas med hjälp av ett konfokalmikroskop för Bodipy (5A) och ett epifluorescensmikroskop för muskelfibertyper (5B). Bilder analyseras i Fiji genom att tillämpa en tröskel och kvantifiera partiklar (6A) för att erhålla antalet, medelstorleken, densiteten och procentandelen av den totala ytan som upptas av LDs (7) eller räkningsceller (6B) för att erhålla procentandelen fibrer av varje typ i sektionen (7). Förkortningar: LDs = lipiddroppar; EDL = extensor digitorum longus; MyHC = myosin tunga kedjeisoformer. Klicka här för att se en större version av denna siffra.
